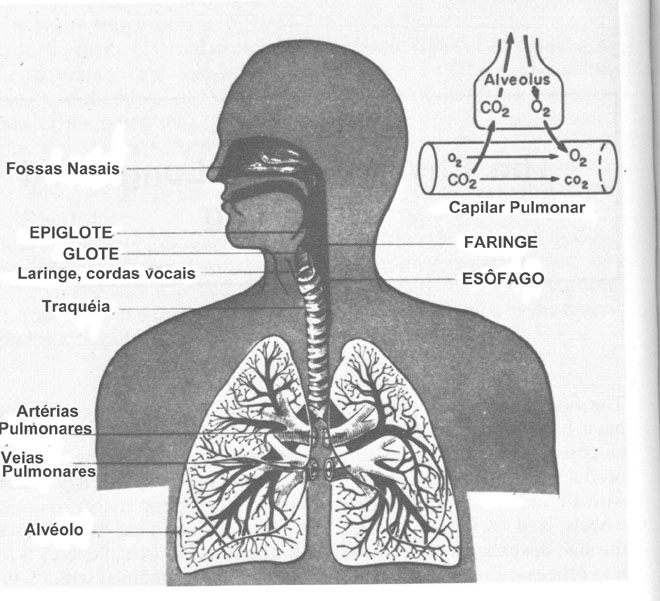
O corpo humano é sob muitos aspectos uma máquina – uma máquina muito interessante. Ele deve ter:
- uma fonte de energia,
- um método de converter a energia nas formas elétrica e mecânica, e
- um modo de expelir os subprodutos.
Num automóvel - a fonte de energia é a gasolina; ela é combinada com o ar e queimada nos cilindros para produzir energia cinética para acionar as rodas, e seu subproduto de gases nocivos e calor são expelidos através do escapamento e radiador.
No corpo humano - a fonte de energia é o alimento; ele é processado no sistema digestivo e é então combinado com O2 nas células do corpo para liberar energia. Seus subprodutos são expelidos de 4 formas:
as
componentes não digestivas são eliminadas como fezes (liberando somente uma
pequena quantidade de gases nocivos),
água e outros subprodutos são expelidos pela
urina,
é expelido via
pulmões a cada dia, e
o calor é dissipado pela superfície do corpo.
A “máquina” humana realmente consiste de bilhões de “máquinas” muito pequenas – as células vivas do corpo. Cada uma destas
“máquinas – miniaturas” deve ser provida com combustível, O2, e um
método de livrar-se dos subprodutos. O sangue e seus vasos (o sistema
cardiovascular) serve como sistema de transporte para estas máquinas. Os
pulmões (o sistema pulmonar) serve como fornecedor de O2 e expelidor
do principal subproduto – CO2. O sangue leva o O2 aos
tecidos e remove o CO2 dos tecidos; ele deve entrar em estreito
contato com o ar nos pulmões a fim de trocar sua carga de CO2 por
uma carga fresca de O2 (Fig.1). Os detalhes deste processo são
discutidos na seção.2.
Por causa
da estreita cooperação e interação entre os sistemas pulmonar e cardiovascular,
as ações de um sistema freqüentemente afetam o outro. Por exemplo, durante a
respiração a pressão nas veias principais do peito afetam o retorno do sangue
ao coração. Freqüentemente uma doença dos pulmões produzirá sintomas no coração
e vice-versa
Figura 7.1 Os canais principais para o ar e sangue no sistema respiratório. O diagrama ilustra esquematicamente as trocas de O2 e CO2 entre o ar nos alvéolos e o sangue. (De Guyton, A.C., Function of the Human Body, 3rd ed. , Ó W.B. Saunders Company, Philadelphia, 1969, p. 222, Reprinted by permission.)
Os pulmões realizam outras funções fisiológicas além das trocas de O2 e CO2. Uma função principal é manter o pH (acidez) do sangue constante. Os pulmões desempenham um papel secundário nas trocas de calor (ver Capítulo 5) e o balanço fluido do corpo por aquecimento e mistura do ar que respiramos (inspiramos). Nosso mecanismo de respiração produz um fluxo controlado de ar para falarmos, tossirmos, espirrarmos, suspirarmos, soluçarmos, sorrirmos, fungarmos e bocejarmos. Em adição, bloqueando a passagem de ar gera aumentos de pressão para defecar e vomitar.
Uma importante função do aparelho respiratório é a produção da voz. As formas de respiração são marcadamente diferentes durante a conversação. Desde que a voz é produzida por uma saída controlada de ar dos pulmões, uma pessoa inala rapidamente e muito profundamente antes de falar afim de ter mais tempo para produzir sons vocais. O tempo de inalação é tipicamente menos de 20% do ciclo da respiração, e a quantidade inalada é geralmente mais que duas vezes o volume usual. Uma cantora (especialmente uma cantora de ópera) inala mesmo mais ar num curto período de tempo para posteriormente minimizar as partes inalatórias do ciclo. A resistência aérea produzida pelas cordas vocais causam um aumento na pressão relativamente grande na traquéia. Assim, o trabalho envolvido no falar e no cantar é consideravelmente maior que o trabalho da respiração normal. Entretanto, relativamente pouco do aumento de trabalho participa da energia sonora.
A voz tipicamente tem uma potência menor que 1 mW. A produção da voz será discutida posteriormente.
Nós respiramos cerca de 6 litros de ar por minuto. (Isto é também cerca do volume de sangue que o coração bombeia cada minuto). A respiração do homem em repouso é cerca de 12 vezes por minuto, a mulher respira cerca de 20 vezes por minuto, e as crianças respiram cerca de 60 vezes por minuto. Discutimos na Seção 7.6 os fatores físicos que afetam a razão de respiração.
O ar que nós inspiramos é cerca de 80% de N2 e 20% de O2. O ar expirado é cerca de 80% de N2, 16% de O2 e 4% de CO2. Nós respiramos cerca de 10 kg (22 lb) de ar a cada dia. Disto o pulmão absorve cerca de 400 litros (~ 0,5 kg) de O2 e libera uma quantidade ligeiramente menor de CO2. Nós também saturamos o ar que nós respiramos com água. Quando respiramos ar seco, nosso ar expirado leva para fora cerca de 0,5 kg de água a cada dia. (Esta mistura pode ser usada para limpar os óculos). No frio alguma desta mistura se condensa e vemos a nossa respiração.
O ar que nós respiramos contém poeira, fumaça, carga de bactérias, gases nocivos e assim por diante, que entram em contato íntimo com o sangue. A grande superfície torcida dos pulmões com uma área superficial de cerca de 80 m2 tem uma exposição maior ao ambiente que qualquer outra parte do corpo incluindo a pele. É talvez surpreendente nós não termos mais doenças dos pulmões. A importância de ar limpo é óbvia.
Cada vez que nós respiramos, cerca de 1022 moléculas de ar entram em nossos pulmões. Lembrem-se que 22,4 litros de ar contém cerca de 6 x 1023 moléculas – número de Avogadro. O número total de moléculas na atmosfera terrestre é cerca de 1044. Assim, tomamos 1/1022 de todo o ar terrestre a cada vez que respiramos; em outras palavras, para cada molécula que respiramos existem 1022 outras mais na atmosfera. A atmosfera terrestre está em constante movimento e sobre um período de séculos tem existido através de mistura de gases. Como um resultado, cada 0,5 litros de ar (1022 moléculas ) contém em média uma molécula que estava presente em qualquer 0,5 litro de ar séculos atrás. Um modo interessante de pensar sobre isto é que na média cada uma de nossa respiração contém uma molécula de ar que foi respirada por Cristo em qualquer uma de suas respirações. É claro, isto é verdadeiro para qualquer pessoa, famosa, não famosa, ou desconhecida que viveu muitos anos atrás.
Problema
1 -
Calcule o número de moléculas de O2 absorvidas pelo corpo numa respiração
típica de 500 cm3.
AS VIAS AÉREAS
Diagrama esquemático mostrando as principais passagens de ar para os pulmões
As principais passagens aéreas para os pulmões estão mostradas na Fig 7.2. O ar entra normalmente no corpo através do nariz onde ele é aquecido (se necessário), filtrado e misturado. As superfícies úmidas e os pêlos no nariz aprisionam particles de poeira, micróbios, e assim por diante. Durante exercícios pesados, tais como jogando, o ar é respirado através da boca e escapam deste sistema de filtro. O ar então passa através da traquéia. A traquéia se divide em duas (bifurca) para fornecer o ar a cada pulmão através dos brônquios. Cada brônquio se divide e subdivide cerca de 15 vezes mais; os bronquíolos terminais resultantes alimentam de ar os milhões de pequenos sacos chamados alvéolos. Os alvéolos, que são como pequenos balões interconectados (Fig. 7.3), têm mais ou menos 0,2 mm de diâmetro (uma folha de papel é aproximadamente 0,1 mm de espessura) e tem paredes de somente 0,4 m m de espessura. Eles expandem-se e contraem-se durante a respiração; eles estão “onde a ação está” na troca de O2 e CO2. Cada alvéolo é envolvido por sangue de modo que O2 possa difundir dos alvéolos para os glóbulos vermelhos e CO2 possa difundir do sangue para o ar no alvéolo. No nascimento os pulmões tem cerca de 30 milhões de alvéolos; aos 8 anos de idade o número de alvéolos cresce para cerca de 300 milhões. Além desta idade o número permanece relativamente constante, mas o alvéolo aumenta de diâmetro. Os alvéolos desempenham um papel tão importante na respiração que discutiremos a física dos alvéolos com mais detalhes na seção 7.5.
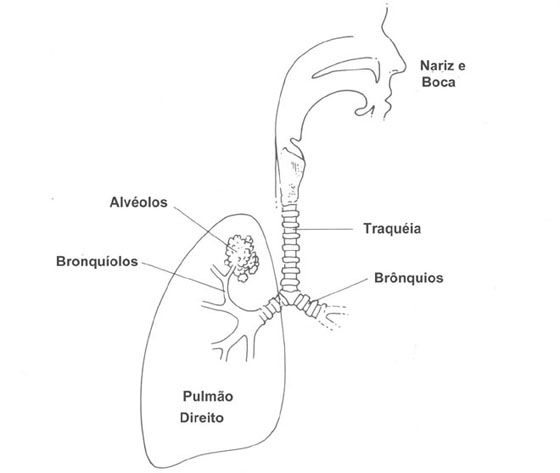
Figura 7.2 - Diagrama esquemático mostrando as passagens principais de ar para os pulmões.
Afim de servir como sistema de transporte para o ar, as vias respiratórias removem as partículas “poeira” que atingem a úmidas vias das várias passagens de ar. O corpo tem dois mecanismos para limpeza das vias respiratórias de partículas estranhas. Os mais corpulentos são removidos pelo tosse. Partículas pequenas são levadas de cima para baixo para a boca por milhões de pequenos pêlos, ou álios. Os álios, que são somente cerca de 0,1 mm de comprimento, tem um movimento ondulatório que move mucosa levando poeira e outras pequenas partículas até os vias respiratórias maiores. Cada um dos álios vibram cerca de 1 000 vezes por minuto. A mucosa move cerca de 1 a 2 cm/min. Você pode pensar nos álios como um sistema de escada rolante para a traquéia. Leva cerca de 30 min para uma partícula de poeira ser retirada dos brônquios e da traquéia para a garganta onde ela é expelida ou engolida.
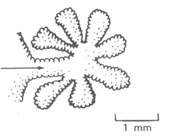
Figura 7.3 – A estrutura do alvéolo
7.2 – COMO O SANGUE E OS PULMÕES INTERAGEM
O principal propósito da respiração é levar um suprimento de O2 fresco para o sangue nos pulmões e desfazer-se do CO2. Nesta seção tentaremos ajudá-lo a entender a física envolvida na troca de gás entre os pulmões e o sangue.
O sangue é bombeado do coração para os pulmões sob pressão relativamente baixa. A pressão sangüínea média de pico na principal artéria pulmonar que leva o sangue aos pulmões é somente cerca de 20 mmHg ou cerca de 15% da pressão na circulação principal do corpo. Os pulmões oferecem pouca resistência ao fluxo de sangue. Na média, cerca de um quinto (~ 1 litro) do sangue que alimenta o corpo está nos pulmões, mas somente cerca de 70 ml daquele sangue está nos capilares dos pulmões obtendo O2 em qualquer instante. Desde que o sangue fica nos capilares pulmonares por menos de 1 segundo, os pulmões devem ser bem projetados para trocas gasosas; os alvéolos dos pulmões tem paredes extremamente finas e são envolvidos pelo sangue no sistema capilar pulmonar. A área da superfície entre o ar e sangue nos pulmões é cerca de 80 m2 (cerca da metade da área de uma quadra de tênis). Se os 70 ml de sangue nos capilares pulmonares forem espalhada sobre uma área superficial de 80 m2 a camada resultante de sangue será somente cerca de 1 mm de espessura, menos do que a espessura de uma única célula de glóbulo vermelho.
Dois processos gerais estão envolvidos nas trocas gasosas nos pulmões:
obtenção do sangue para leito capilar pulmonar (aspersão) e
obtenção de ar para as superfícies alveolares (ventilação).
Se um ou outro processo falha o sangue não será oxigenado apropriadamente.
Existem três tipos de áreas aspersão-ventilação nos pulmões:
áreas com boa ventilação e boa aspersão,
aquelas com boa ventilação e aspersão pobre, e
aquelas com pobre ventilação e boa aspersão.
Num pulmão normal o primeiro tipo ocorre com mais de 90% do volume total. Se o sangue que flui para uma parte de um pulmão é bloqueado por um coágulo (uma embolia pulmonar) aquele volume terá aspersão pobre. Se as passagens aéreas nos pulmões estão obstruídas como na pneumonia, a área envolvida terá ventilação pobre. Muitas doenças pulmonares causam reduções na aspersão ou ventilação.
A transferência de O2 e CO2 para dentro e para fora do sangue é controlada pela lei física da difusão. Todas as
moléculas estão continuamente em movimento. Nos gases e nos líquidos, e numa
certa extensão mesmo nos sólidos, as moléculas não permanecem numa determinada
localização. Por exemplo, se você pode identificar um grupo de moléculas numa
sala (p. ex., de uma gota de perfume) em poucos minutos você encontrará que
estas moléculas moveram-se (difundiram-se) pela sala. Moléculas de um tipo
particular difundem-se de uma região de alta concentração para uma região de
concentração mais baixa até que a concentração seja uniforme. Nos pulmões
estamos interessados na difusão de gases e líquidos. As moléculas num gás na
temperatura da sala movem-se aproximadamente com a velocidade do som. Cada
molécula colide cerca de 1010 vezes em cada segundo com as moléculas
vizinhas, peregrinando no processo de uma maneira aleatória. A distância D mais
provável que uma molécula viajará de sua origem após N colisões é D = l ![]() , onde l é o caminho livre médio, ou a distância média entre as
colisões. No ar l
é cerca de 10-7 m; no tecido l é cerca de 10-11
m (ver Exemplo 7.1).
, onde l é o caminho livre médio, ou a distância média entre as
colisões. No ar l
é cerca de 10-7 m; no tecido l é cerca de 10-11
m (ver Exemplo 7.1).
EXEMPLO 7.1
Qual é o valor típico de D no ar e no tecido para uma molécula de O2 após 1 segundo se N = 1010 no ar e N = 1012 no tecido?
No ar D ~ 10-7 (1010)1/2 = 10-2 m
No tecido D ~ 10-11 (1012)1/2 = 10-5 m.
A difusão
depende da velocidade das moléculas; ela é mais rápida se as moléculas são
leves e ela cresce com a temperatura. Desde que N é proporcional ao tempo de
difusão Dt
(i.é, N a
Dt),
podemos escrever que D a ![]() ou Dt a D2.
Se D = 10 mm após 1 segundo, a molécula levará 100 segundos, em média, para
difundir 100 mm. Nos pulmões a distância a ser viajada no ar é usualmente uma
pequena fração de um milímetro, e a difusão toma lugar numa fração de um
segundo. A difusão de O2 e CO2 no tecido é cerca de 10
000 vezes mais lenta que no ar, mas a espessura do0tecido pelo qual a molécula
deve difundir-se nos pulmões é muito pequena (~0,4 mm ) e a difusëo através di
parede alveolar leva muito menos que 1 segundo. Discutimos mais sobre a difusão
nos tecidos em outro Apêndice.
ou Dt a D2.
Se D = 10 mm após 1 segundo, a molécula levará 100 segundos, em média, para
difundir 100 mm. Nos pulmões a distância a ser viajada no ar é usualmente uma
pequena fração de um milímetro, e a difusão toma lugar numa fração de um
segundo. A difusão de O2 e CO2 no tecido é cerca de 10
000 vezes mais lenta que no ar, mas a espessura do0tecido pelo qual a molécula
deve difundir-se nos pulmões é muito pequena (~0,4 mm ) e a difusëo através di
parede alveolar leva muito menos que 1 segundo. Discutimos mais sobre a difusão
nos tecidos em outro Apêndice.
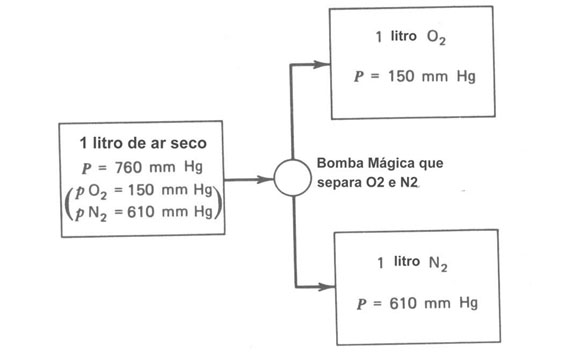
Figura 7.4. Uma ilustração esquemática da lei de Dalton das pressões parciais. Um litro de ar a uma pressão de760 mmHg pode ser pensado como uma mistura de 1 litro de O2 na pressão de 150 mmHg e 1 litro de N2 na pressão de 610 mmHg
Para entender o comportamento dos gases nos pulmões é necessário revisar a lei de Dalton das pressões parciais. Esta lei diz que se você tem uma mistura de vários gases, cada gás faz a sua própria contribuição à pressão total imaginada como se ele fosse o único. Consideremos um recipiente fechado de ar seco à pressão atmosférica (760 mmHg). Se você removeu do recipiente todas as moléculas exceto O2, a pressão cairá para cerca de 150 mmHg (i.é, 20% de 760 mmHg). Esta é a pressão parcial do oxigênio pO2. Se somente as moléculas de N2 forem deixadas, a pressão será cerca de 80% de 760 mmHg ou cerca de 610 mmHg. A Figura 7.4 mostra esquematicamente este experimento imaginário. A pressão parcial do vapor d’água no ar depende da umidade. No ar de uma sala típica, a pressão parcial do vapor d’água é de 15 a 20 mmHg; nos pulmões a 37ºC e 100% de umidade relativa a pressão parcial do vapor d’água é 47 mmHw.
Consideremos o que acontece num recipiente fechado com sangue e O2. Algumas moléculas de O2 colidiróo com o sangue e serão dissolvidas. Após, um tempo o número de molésulas de O2 que estão(escapando do sangue a cada segundo é o mesmo número das que(estão entraodo nele/ O sangue tem então uma pO2 igual aquela do O2 em contato com ele. Se a pO2 na fase gasosa é eobrada, a quantidade de O2 dissolvida no sangue também será dobrada. Esta proporcionalidade é a lei de Henry da solubilidade dos!gases.
A quantidade de gás disóolvida no sangue varia castante de um gás para o outro. O oxigênio não é muito solúvel oo sangue ou água. À temperaôura(do corpo, um litro de plasma saogüíneo à pO3 te 100 mmHg manterá somente cerca de 2,5 cm3 de O2 a temperatura e pressão normal (C.N.T.P.). Em pO2 de 40 mmHg manterá cerca de 25 cm3 de CO2 na solução. Se o corpo teve de depender de O2 dissolvido no plasma para suprir O2 para as células, o coração terá que bombear 140 litros de sangue por minuto no repouso, em vez dos 6 litros/minuto que ele realmente bombeia. Como discutimos brevemente, existe um método mais eficiente de transporte de O2 e CO2 que envolve os glóbulos vermelhos.
As diferentes solubilidades de O2 e CO2 no tecido afetam o transporte destes gases através da parede alveolar. Uma molécula de O2 difunde-se mais rapidamente que uma molécula de CO2 devido a sua massa ser menor. Entretanto, por causa do maior número de moléculas de CO2 na solução, o transporte de CO2 é mais eficiente que o transporte de O2. Se uma doença torna a parede alveolar mais espessa, o transporte de O2 é mais prejudicado que o transporte de CO2.
A mistura de gases nos alvéolos não é a mesma que a mistura de gases no ar ordinário. Os pulmões não são esvaziados durante a expiração. Durante a respiração normal os pulmões retém cerca de 30% do seu volume no final de cada expiração. Isto é chamado “CAPACIDADE RESIDUAL FUNCIONAL” (F.R.C.). Em cada respiração cerca de 500 cm3 de ar fresco (pO2 de 150 mmHg) mistura-se com cerca de 2 000 cm3 de ar viciado nos pulmões para resultar no ar alveolar com uma pO2 de cerca de 100 mmHg. O pCO2 nos alvéolos é cerca de 40 mmHg. O ar expirado inclui cerca de 150 cm3 de ar relativamente fresco da traquéia que não estava em contato com as superfícies alveolares, assim o ar expirado tem uma pO2 ligeiramente mais alta e uma pCO2 mais baixa que o ar alveolar (Tabela 7.1). A razão da saída do CO2 pela entrada de O2 é chamada RAZÃO DE TROCA RESPIRATÓRIA ou QUOCIENTE RESPIRATÓRIO R (ver Capítulo 5). Geralmente R é ligeiramente menor que 1.
As porcentagens e Pressões Parciais de O2 e CO2 no Ar Inspirado, Alveolar e Expirado
O nitrogênio do ar não desempenha qualquer papel conhecido nas funções do corpo. Ele é dissolvido no sangue na sua pressão parcial. Um mergulhador no mar respira ar numa pressão muito mais alta submerso do que quando ele está no nível do mar; a pressão parcial aumentada de N2 faz com que mais N2 seja dissolvido no seu sangue e tecidos. Se o mergulhador vem à superfície muito rapidamente alguns dos N2 formam bolhas nas suas juntas causando os sérios problemas de cãimbras (ver Capítulo 6).
Durante a respiração normal o suprimento de ar fresco não entra nos alvéolos que estão ainda cheios de ar viciado da respiração anterior. Por causa da sua maior concentração, o O2 fresco rapidamente difunde pelo ar viciado até atingir a superfície dos alvéolos. O O2 é dissolvido na mistura parede alveolar e difunde por ela até o capilar sangüíneo até que a pO2 no sangue seja igual aquela dos alvéolos. Este processo leva menos do que 0,5 segundos (Fig. 7.5). Entretanto, o CO2 no sangue difunde-se mesmo mais rapidamente no gás dos alvéolos até que a pCO2 no sangue seja a mesma que no gás alveolar.
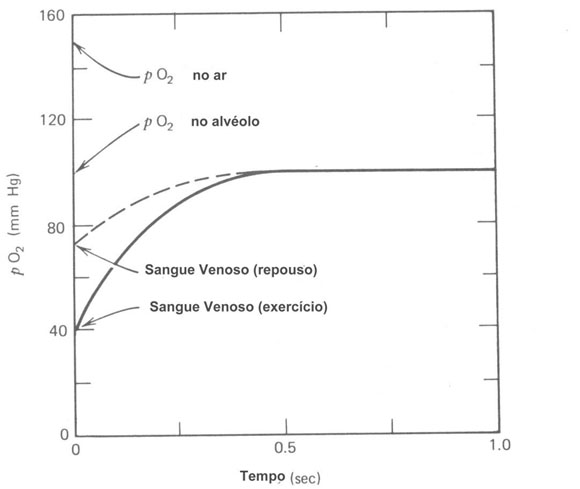
Figura 7.5– O pO2 do sangue num capilar pulmonar aumenta rapidamente o nível da pO2 nos alvéolos quando os glóbulos vermelhos movem pelo capilar (linha tracejada). Mesmo durante exercícios pesados (linhas sólidas) os glóbulos vermelhos são rapidamente reabastecidos de O2.
Como mencionado anteriormente, o sangue pode carregar muito pouco O2 na solução. A maioria do O2 para as células é carregado na combinação química com a hemoglobina (Hb) nos glóbulos vermelhos. Um litro de sangue pode carregar cerca de 200 cm3 de O2 nas C.N.T.P., por este meio enquanto ele pode carregar somente 2,5 cm3 de O2 em solução. Desde que a maioria do O2 não está em solução, a lei de difusão á alterada. O O2 combinará com ou separará da Hb de uma maneira que depende da curva de dissociação (Fig. 7.6). A Hb que deixa os pulmões está cerca de 97% saturada com O2 numa pO2 de cerca de 100 mmHg. O pO2 tem que cair para cerca de 50% antes da carga de O2 do sangue é notavelmente reduzida.
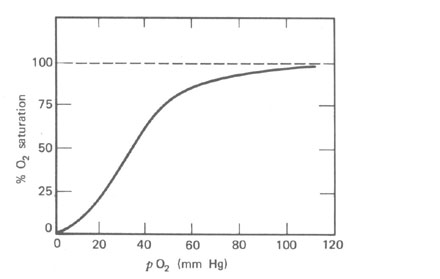
Figura 7.6 - A porcentagem de saturação de O2 do sangue como uma função de pO2 nos alvéolos. A 100% de saturação 1 litro de sangue pode transportar 200 ml de O2 nas CNTP. Esta curva é afetada pela temperatura, ao pCO2, e o pH.
Quando o sangue atinge as células e seu baixo pO2 ambiente, o O2 é dissociado do Hb e difunde na célula. Nem todo o O2 deixa a Hb; a quantidade deixada depende do pO2 dos tecidos. Sob condições de repouso o sangue venoso retorna ao coração com cerca de 75% da sua carga de O2. O O2 é retido no sangue porque não é necessário nos tecidos. Durante trabalhos físicos pesados ou exercícios a situação nos músculos ativos variam drasticamente. O pO2 nos músculos em funcionamento cai rapidamente fazendo mais O2 ser dissociados da Hb e difundir para o interior dos músculos. Em adição o corpo pode aumentar o fluxo sangüíneo para o trabalho muscular por um fator de 3. Os músculos trabalhando podem obter 10 vezes mais O2 que eles consomem no repouso. Para pessoas normais o fator limitante no exercício não é a quantidade de sangue bombeado pelo coração por minuto (saída cardíaca) ou a quantidade de O2 fornecida ao sangue pelos pulmões, mas a velocidade em que O2 é transferido aos músculos trabalhando.
A dissociação de O2 da Hb também depende do pO2, do pH (acidez) e da temperatura. Durante os exercícios o pCO2, a acidez e a temperatura nos músculos trabalhando aumentam todos; este aumento desloca todas as curvas da Fig. 7.6 para a direita e permite a Hb dar mais do seu O2. Todos estes fatores aumentam assim o O2 para os músculos trabalharem. Nos pulmões a diminuição de pCO2 devido a rápida respiração permite Hb agarrar mais O2.
Dióxido de carbono não é transportado dos tecidos por simples difusão. (Se você está interessado nos detalhes ver bibliografia no final deste capítulo). A maioria do CO2 permanece no sangue após ele ter deixado os pulmões (pCO2 ~ 40 mmHg). O nível de CO2 no sangue é mantido bem constante pela razão de respiração. Respiração excessivamente rápida (hiperventilação) pode abaixar o pCO2 do sangue (hipocapnia); isto provoca distúrbios mentais e desmaios.
No envenenamento por monóxido de carbono (CO) moléculas são grudadas muito seguramente à Hb nos lugares que normalmente seriam ocupados por O2. Elas grudam cerca de 250 vezes mais que o O2 e não se dissociam facilmente nos tecidos. Em adição usando os lugares normalmente usados para transportar O2, o CO impede a liberação de O2 da Hb, todavia uma pequena quantidade de CO possa seriamente reduzir o O2 nos tecidos. Fumantes de cigarros respiram em cerca de 250 cm3 de CO em cada pacote, e também é geralmente inalado por pessoas viajando em tráfego pesado. Monóxido de carbono pode causar a morte por desnutrição dos tecidos de O2. Normalmente o O2 dissolvido no sangue é de não significado, mas se uma vítima de CO é colocada numa câmara hiperbárica de O2 com uma pressão absoluta de 3 atm de O2 puro, o pO2 aumenta por um fator de 15. O O2 dissolvido no sangue pode então suprir as necessidades mínimas do corpo (ver Capítulo 6). Esta terapia não pode ser mantida por muito tempo porque o envenenamento por O2 pode resultar. Uso continuado de 1 atm de O2 puro pode causar edema dos tecidos dos pulmões, que reduz o O2 no sangue e ironicamente resulta na morte de uma falta de O2 (anóxia = deficiência de oxigênio nos tecidos). Níveis de proteção de pO2 no “ar” são aqueles abaixo de 0,5 atm (~ 380 mmHg pO2) (ver Fig. 6.8).
7.3 MEDIDAS DOS VOLUMES PULMONARES
Um instrumento relativamente simples, o espirômetro (Fig. 7.7), é usado para medir o fluxo de ar para dentro e para fora dos pulmões e registrar num gráfico o volume versus o tempo. A Figura 7.8 mostra um registro típico para um adulto sob várias condições de respiração. Durante a respiração normal no repouso nós inalamos cerca de 500 cm3 de ar em cada respiração. Isto é referido como volume tidal (maré) no repouso. No começo e no final de uma respiração normal existe reserva considerável. No final de uma inspiração normal é possível com algum esforço promover o enchimento dos seus pulmões com ar. O ar adicional tomado é chamado volume de reserva inspiratório. Similarmente, no final de uma expiração normal você pode forçar mais ar para fora dos seus pulmões . Este ar adicional expirado é chamado volume de reserva expiratório. O ar restante nos pulmões após uma expiração normal é chamado capacidade residual funcional (FRC). É este ar velho que mistura com o ar fresco na nova respiração. Durante exercícios pesados, o volume tidal é consideravelmente maior. Você tem uma clara idéia da sua capacidade pulmonar se você tem mesmo soprar um saco de papel ou um colchão de ar. Se uma pessoa respira tão profundamente quanto pode (a em Fig. 7.8) e então exala tanto quanto pode (b em Fig. 7.8), o volume de ar exalado é a sua capacidade vital. Entretanto, seus pulmões ainda conterá algum ar – o volume residual, que é cerca de 1 litro para um adulto. O volume residual pode ser determinado tendo submetido a respiração num volume conhecido de um gás inerte tal como o hélio e então medindo a fração do hélio no gás expirado. Desde que o hélio e o ar misturar-se-ão durante uma única respiração, esta técnica de diluição é muito acurada.
Uma grande quantidade de testes clínicos podem ser feitos com o espirômetro. A quantidade de ar respirado em 1 minuto é chamado volume respiratório por minuto. O máximo volume de ar que pode ser respirado em 15 segundos é chamado ventilação voluntária máxima e é uma quantidade clínica útil . A máxima razão de expiração após uma inspiração máxima é um teste útil para enfisema e outras doenças das vias respiratórias obstruídas. Em alguns casos a razão de fluxo decresce com o esforço expiratório excessivo. Uma pessoa normal pode expirar cerca de 70% de sua capacidade vital em 0,5 segundos, 85% em 1,0 segundos, 94% em 2,0 segundos, e 97% em 3,0 segundos. Os picos normais da razão de fluxo são 350 a 500 litros/min. A velocidade do ar expirado pode ser impressiva; se uma pessoa engole ou espirra forte sem cobri sua boca, a velocidade do ar na sua traquéia pode atingir Mach 1 – a velocidade do som no ar! Esta velocidade alta pode causar colapso parcial das vias respiratórias por causa do efeito de Bernoulli. Engolindo para expulsar um objeto estranho este colapso parcial aumenta a velocidade do ar e aumenta a força no objeto estranho.
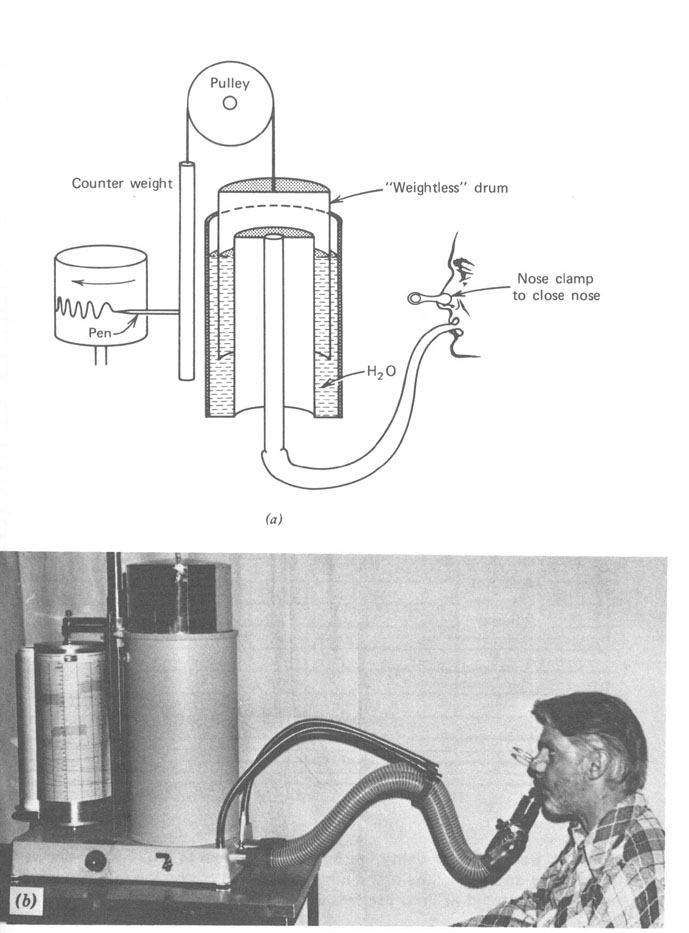
Figura 7.7 - O espirômetro é usado para medir várias quantidades de funçòes pulmonares. O fluxo de ar para dentro e para fora dos pulmões é registrado num cartão giratório. (a) Uma seção transversal do espirômetro mostrando como a água é usada como um selo de ar para manter o ar dentro do tambor contrabalançado. (b) Um dos autores (JRC) produzindo o gráfico mostrado na figura 7.8. O nariz tampado força todo o ar fluir através da boca
Nem todo o ar inspirado adiciona O2 ao sangue. O volume da traquéia e brônquios é chamado espaço anatômico morto já que o ar neste espaço não é exposto ao sangue nos capilares pulmonares. O espaço anatômico morto é cerca de 150 cm3. Em adição , em algumas doenças alguns dos capilares alveolares não são aspergidos com sangue e o O2 não é absorvido neste alvéolo. Este volume não usado é chamado fisiológico ou espaço alveolar morto. O ar no espaço alveolar morto não fornece qualquer O2 ao corpo. O ar no espaço anatômico morto após uma expiração é levado de volta para os pulmões durante a próxima inspiração. Se você aumentar o seu espaço morto respirando através de um tubo , você reciclará mais de sua própria respiração. Se o tubo tem um volume igual à sua capacidade vital você obviamente não obterá ar fresco e sufocará.
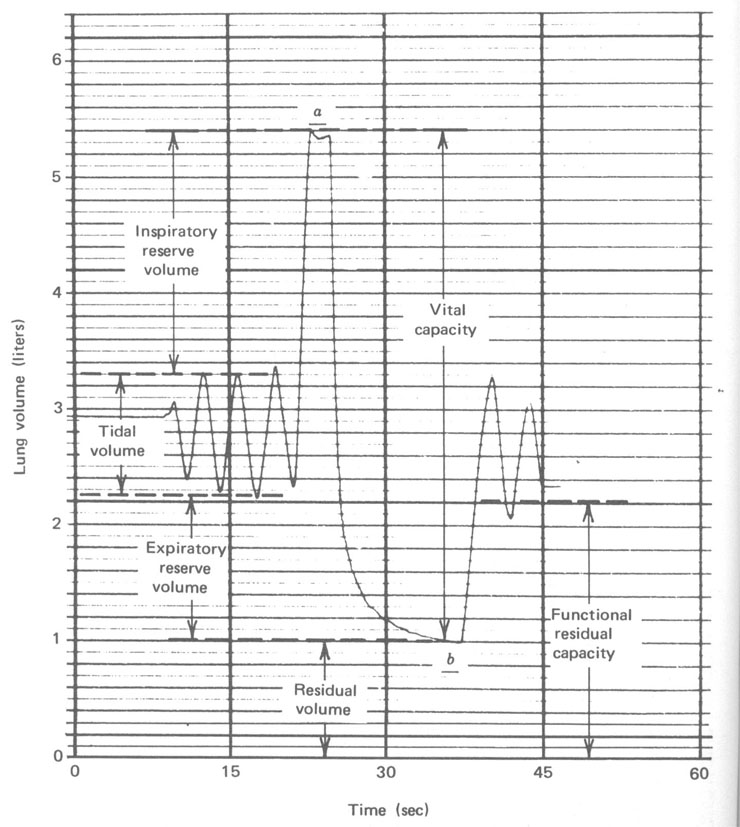
Figura 7.8 - Um traçado feito usando o aparelho mostrado na Figura 7.7b. Ele mostra os vários volumes e capacidades dos pulmões. Note que durante a expiração máxima o ar fluindo para fora é rápido no primeiro momento; os últimos 5% leva mais tempo que os primeiros 95%.
4 RELAÇÕES DA PRESSÃO-FLUXO DE AR-VOLUME DOS PULMÕES.
As relações de pressão, fluxo de ar, e volume dos pulmões durante a respiração tidal para um sujeito normal e para um paciente com uma estreita via aérea estão mostradas na Fig.7.9. A diferença de pressão necessária para fazer o ar fluir para dentro e para fora dos pulmões de um indivíduo saudável é muito pequena. Note que a diferença de pressão (Fig. 7.9a) é somente uns poucos centímetros de água para um indivíduo normal. A Figura 7.9b mostra a razão do fluxo de ar para dentro e para fora dos pulmões em litros por minutos, e a Fig. 7.9c mostra o volume dos pulmões durante o ciclo da respiração.
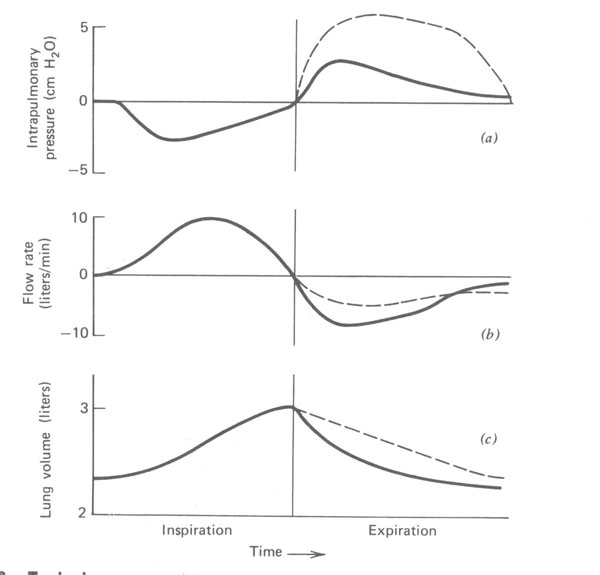
Figura 7.9 - Pressões típicas (a), razão de fluxo (b), e volumes pulmonares (c) durante uma respiração tranquila para um indivíduo normal (linha sólida) e um paciente com uma via respiratória estreita (linha tracejada). Note o acréscimo de pressão e o decréscimo da razão de fluxo durante a expiração no paciente com as vias respiratórias estreitadas
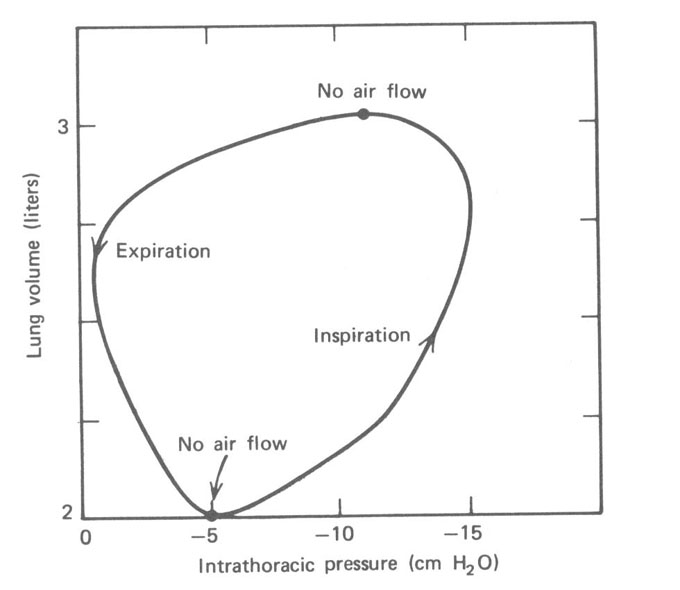
Como o esôfago passa através do peito, ele reflete a pressão entre os pulmões e a parede do peito (intrapleural ou espaço intratorácico). É possível medir a pressão no esôfago com uma pressão gauge. Esta pressão é normalmente negativa (~ -10 mmHg) devido à elasticidade dos pulmões (ver Seção 7.6). Na Fig. 7.10, a pressão intratorácica (medida no esôfago) é plotada versus o volume tidal dos pulmões durante a respiração. A Figura 7.11 mostra as curvas pressão-volume para três diferentes razão de respiração – lenta, moderada e rápida.
Os pulmões e o peito estão normalmente acoplados. O comportamento do sistema é o resultado da combinação das características físicas dos dois. A Figura 7.12 mostra as curvas de volume versus pressão para as paredes do peito e pulmões separadamente e para os dois juntos. O volume é dado como uma porcentagem da capacidade vital. Se a parede do peito estivesse livre de sua interação com os pulmões ela teria um volume de cerca de dois terços da capacidade vital total. Os pulmões por si só murchariam e não teriam volume de ar. Juntos os pulmões e a parede do peito vão a um volume de relaxação (FRC) de cerca de 30% de capacidade vital.
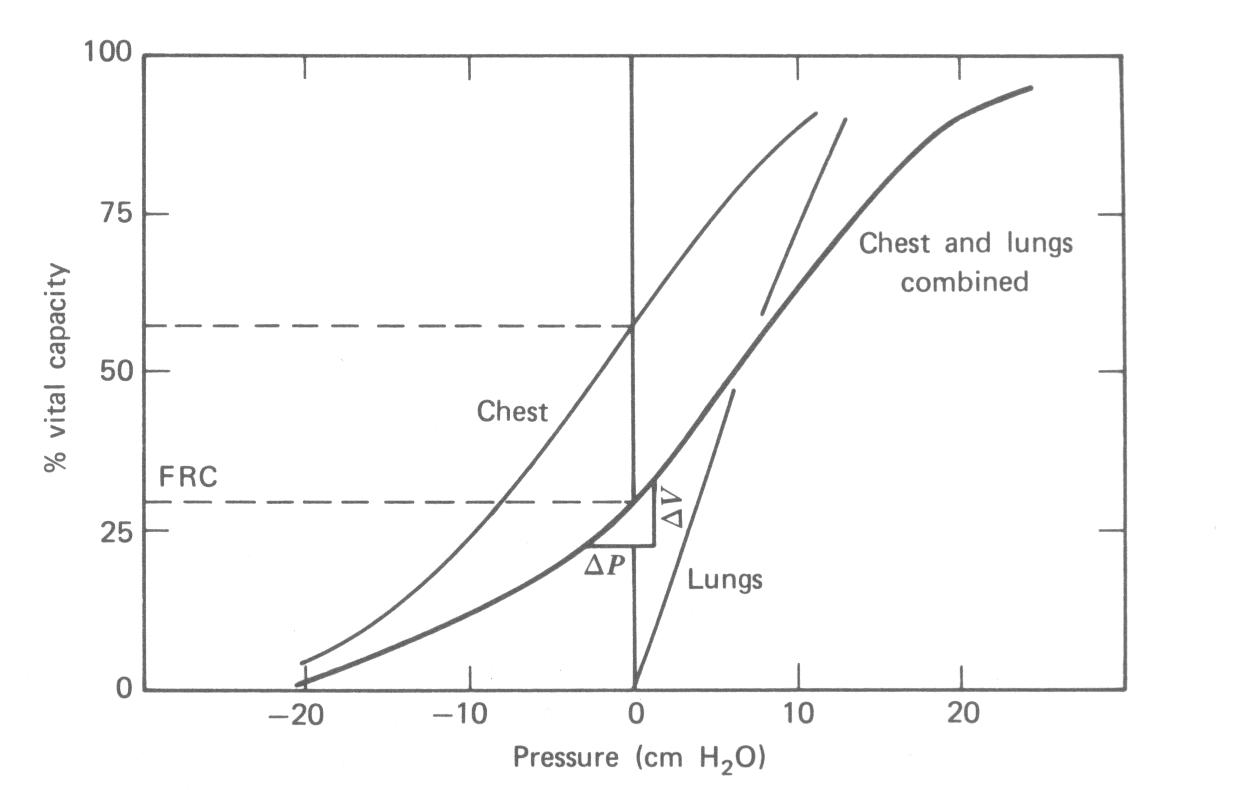
A curva combinada na Fig. 7.12 mostra a relação pressão-volume obtida enchendo os pulmões para conhecer as porcentagens da capacidade vital. A pressão é medida na boca (e pulmões) com o nariz e boca fechados e os músculos da respiração relaxados. Por exemplo, em cerca de 60% de capacidade vital, a pressão de relaxação é 10 cm H2O. Como a parede do peito está em equilíbrio neste volume, esta pressão é produzida pelas propriedades elásticas dos pulmões. Quando a mesma medida de relaxação são feitas após uma exalação forçada os valores de pressão negativa da Fig. 7.12 são obtidos.
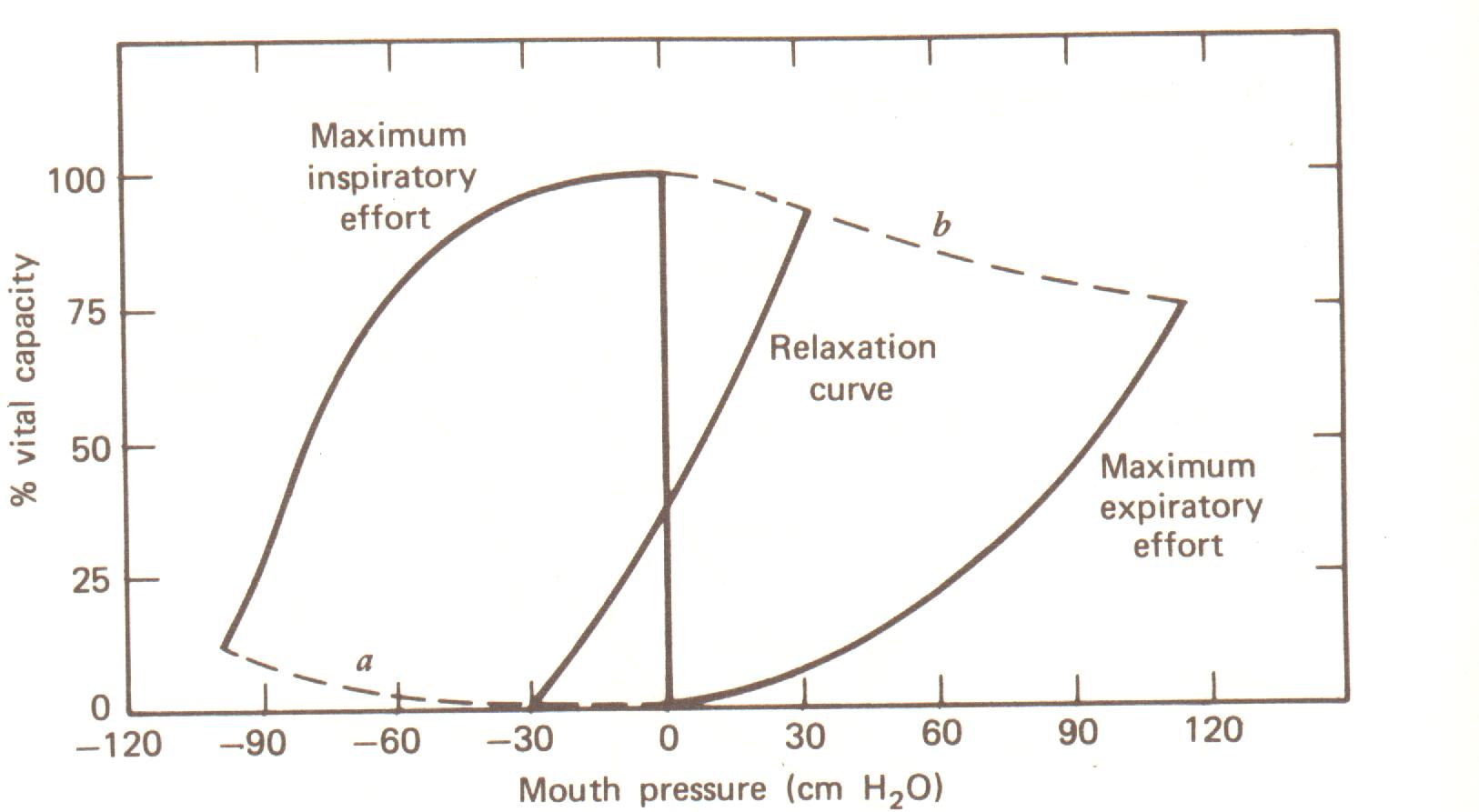
A curva de pressão de relaxação é novamente plotada como uma função da capacidade vital na Fig. 7.13. Ainda, duas outras curvas relacionadas são mostradas. Todas estas curvas são medidas na boca com o nariz e boca fechados. Exalando com uma força maior dá a curva do esforço expiratório máximo. Inalando com o máximo esforço dá a curva do máximo esforço inspiratório. Esforço expiratório forçado após uma inspiração máxima (100% da capacidade vital) comprime o gás de acordo com a lei de Boyle, PV = constante. As linhas tracejadas a e b mostram as curvas teóricas para a relação pressão-volume de um gás ideal (PV = constante) a 0% e 100% de capacidade vital.
Complacência é uma característica física importante dos pulmões. Complacência é a variação no volume produzida por uma pequena variação na pressão, isto é, DV/DP (ver Fig. 7.12). Complacência é usualmente dada em litros por centímetro de água. Complacência nos adultos normais está no intervalo de 0,18 a 0,27 litros/cm H2O. Ela é geralmente cerca de 25% maior no homem acima de 60 anos de idade do no homem jovem. Existe pouca variação com a idade na mulher.
Uma fibrose pulmonar tem uma pequena variação no volume para uma grande variação da pressão e assim ela tem uma baixa complacência. Um pulmão frouxo tem uma grande variação no volume para uma pequena variação na pressão e tem uma grande complacência. Crianças com síndrome de embaraço respiratório (ver Seção 7.5) tem pulmões com baixa complacência. Em algumas doenças, tais como enfisema, a complacência aumenta (ver Seção 7.9).
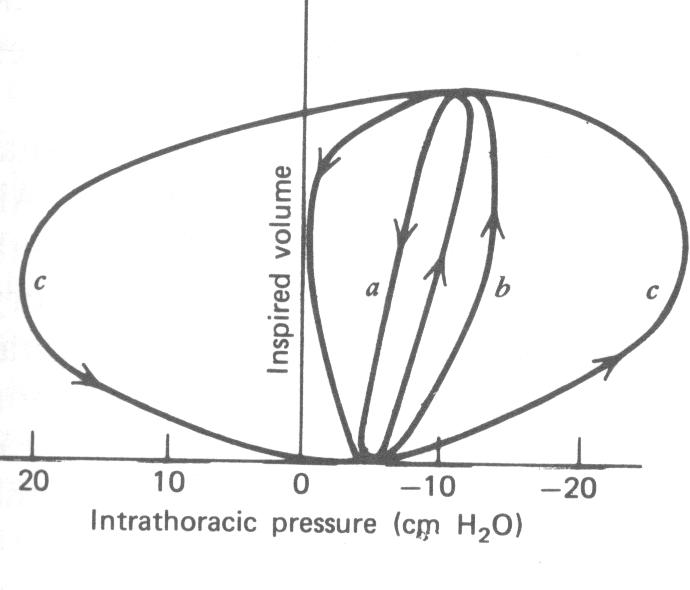
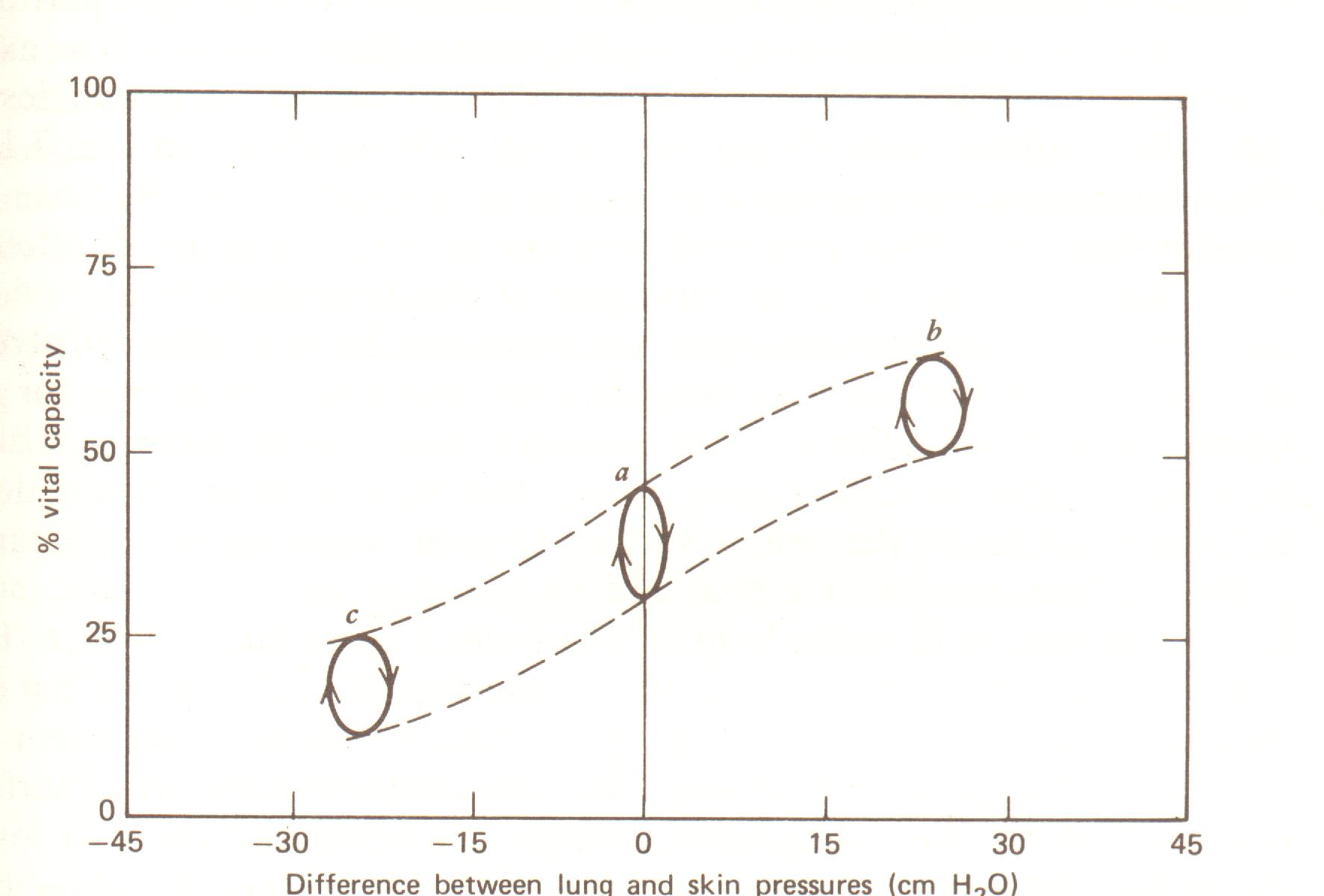
Durante a respiração tidal, a curva P-V forma uma linha fechada como aquela mostrada na Fig. 7.14. O ciclo flui no sentido horário no laço. O laço médio representa uma respiração tidal típica na pressão normal . O laço b representa uma pressão de respiração positiva onde o ar fornecido ao pulmão é cerca de 30 cm H2O maior que a pressão na parede do peito. Pressão de respiração positiva é freqüentemente usada terapeuticamente na ressuscitação e no socorro de doenças obstrutivas das vias respiratórias. Para pressão de respiração positiva os músculos inspiratórios não são usados mas os músculos expiratórios são. O laço c na Fig. 7.14 representa pressão de respiração negativa. Isto pode ocorrer quando uma pessoa está debaixo d’água e respirando através de um tubo para a superfície (através do tubo de respiração para mergulhadores). Neste caso os músculos inspiratórios nunca relaxam completamente.
PROBLEMAS
A FÍSICA DOS ALVÉOLOS.
Os alvéolos são fisicamente como milhões de pequenas bolhas interconectadas. Eles tem uma tendência natural de ficarem menores devido à tensão superficial de um fina camada fluida de revestimento. Este revestimento, chamado surfactante, é necessário para os pulmões funcionarem apropriadamente. A ausência do surfactante nos pulmões de alguma criança recém-nascida, especialmente prematuras, é a causa da síndrome do distúrbio respiratório idiopatico (RDS), algumas vezes chamado doença da membrana hialina. Esta doença contabiliza milhares de mortes infantis a cada ano nos Estados Unidos; ela mata mais crianças que qualquer outra doença .
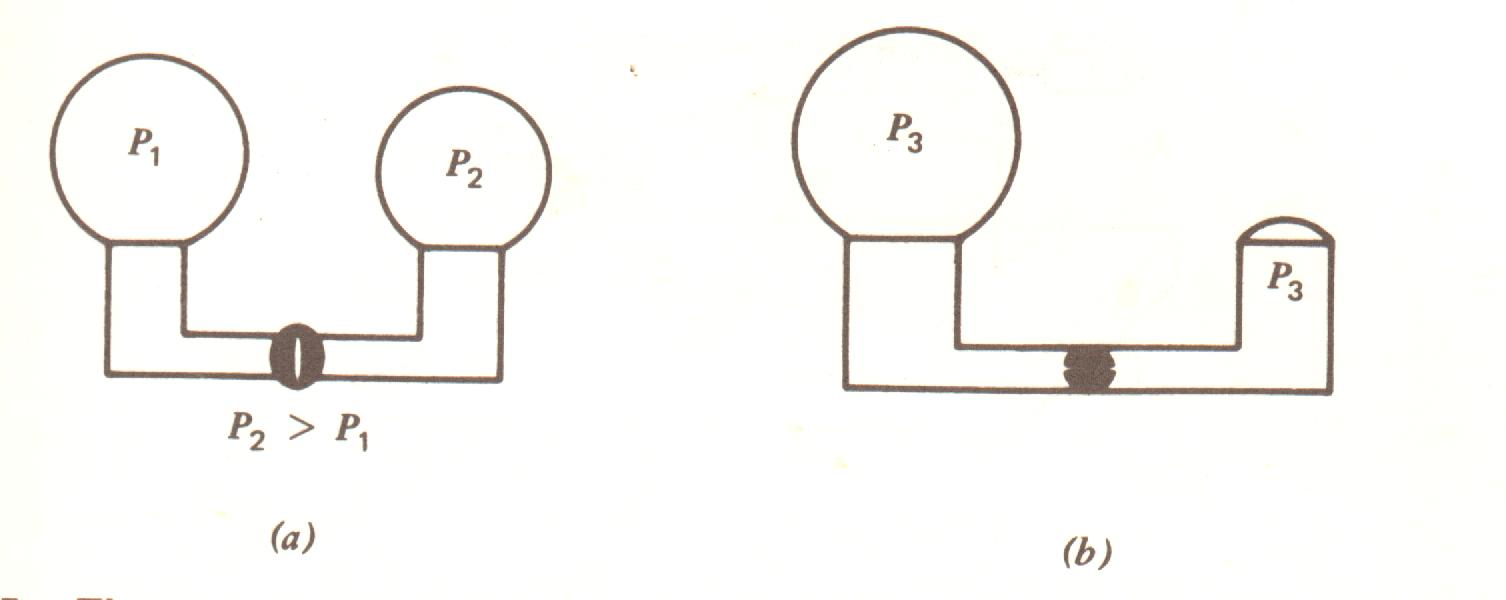
Para entender a física do alvéolo temos de entender a física das bolhas. A pressão dentro de uma bolha é inversamente proporcional ao raio e diretamente proporcional à tensão superficial T. A relação exata é P = 4 T / R, uma forma da lei de Laplace. Consideremos uma bolha de sabão nos extremos de um tubo com uma válvula separando-as como mostrado na Fig. 7.15 a. O que acontece quando a válvula é aberta para conectá-las? Porque a menor tem uma pressão interna maior ela esvaziará o seu ar para o interior da maior até que o raio de curvatura da bolha maior e do restante da bolha menor tornarem-se o mesmo. (Fig. 7.15b). Embora os alvéolos não são exatamente a mesma coisa que bolhas de sabão existe uma tendência do alvéolo menor colapsar. A condição que resulta quando um número considerável colapsam é chamada atelectasia. A razão da maioria dos alvéolos não murcharem está relacionada unicamente às propriedades da tensão superficial da surfactante.
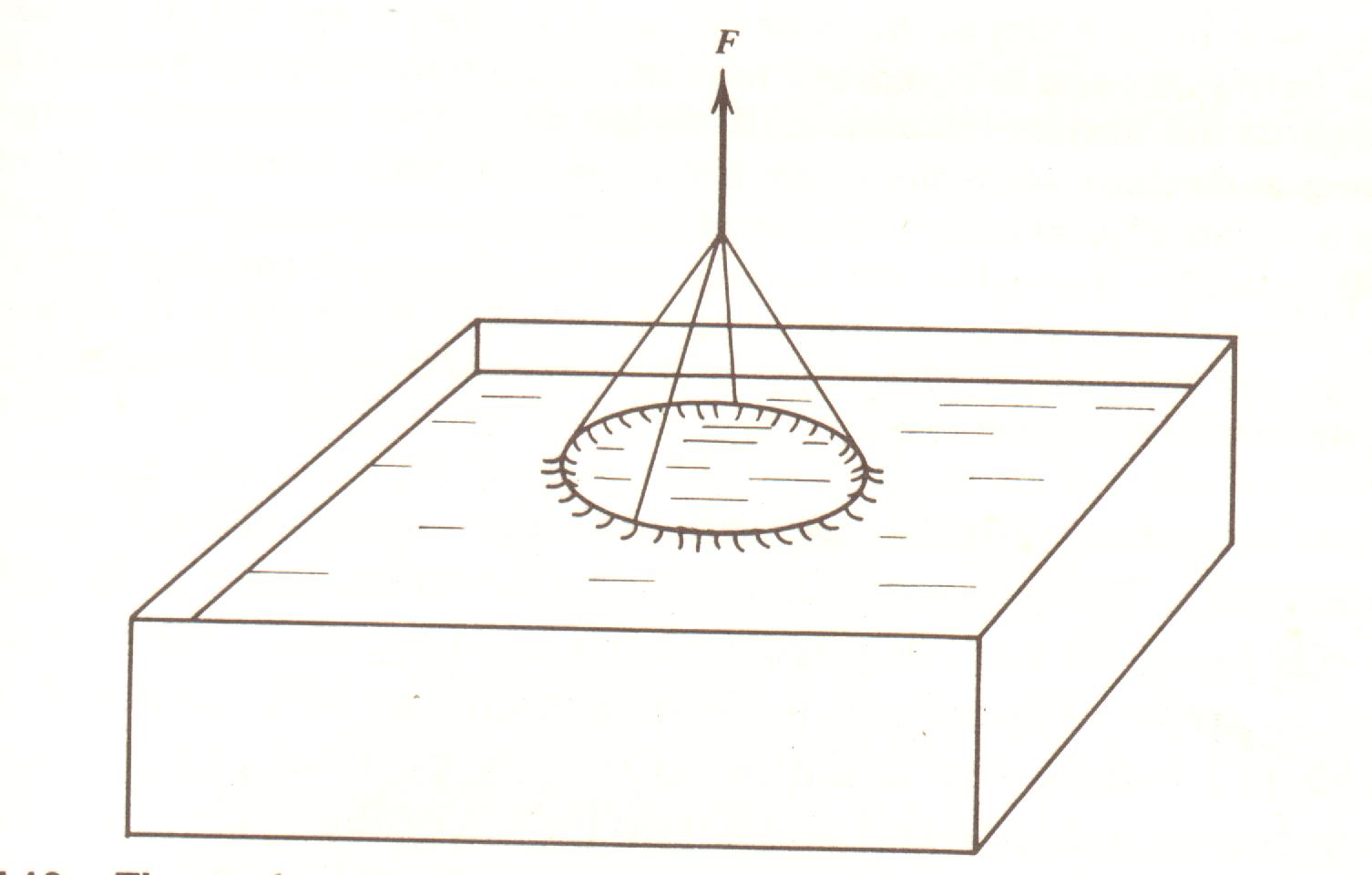
A tensão superficial de um fluido pode ser encontrada medindo-se quanta força é necessária para puxar uma espira de arame de uma superfície líquida clara (Fig. 7.16). A tensão superficial de uma interface água-ar é 72 dinas/cm; aquela de uma interface plasma-ar é cerca de 40 a 50 dinas/cm; aquela da solução de detergente no ar são de 25 a 45 dinas/cm. Uma medida qualitativa da tensão superficial é notar quanto tempo pequenas bolhas de um líquido sobrevivem. Quanto mais baixa a tensão superficial, mais tempo elas resistem. Em 1955 foi notado que bolhas exprimidas dos pulmões eram muito estáveis, resistindo por horas. Foi concluído que elas devem ter uma tensão superficial muito baixa.
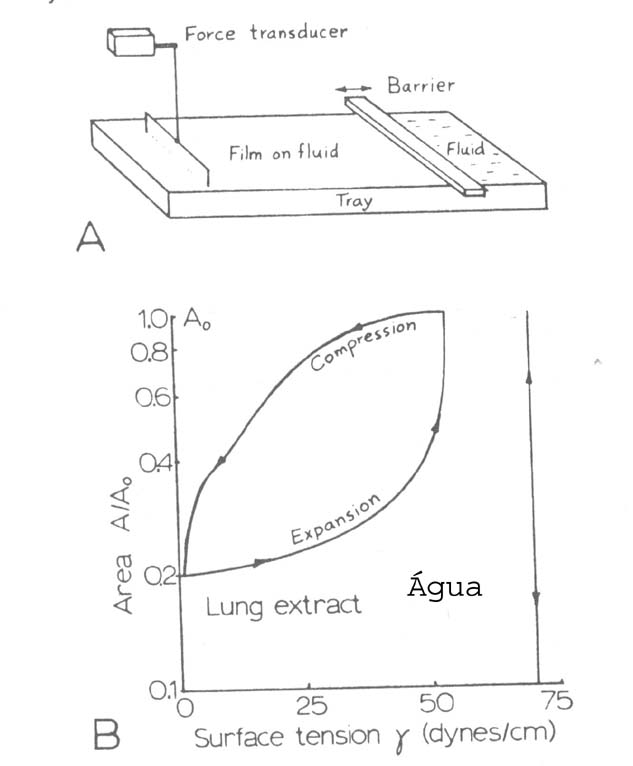
A tensão superficial do surfactante que contornam o alvéolo de um indivíduo saudável faz o papel principal nas funções dos pulmões. A tensão superficial do sufarctante não é constante. A Fig 7.17b mostra a tensão superficial de um filme de extrato pulmonar normal contendo surfactante. Note a grande diminuição de T quando a área diminui. Esta característica faz a tensão superficial do alvéolo diminuir quando o alvéolo diminui de tamanho durante a expiração. Para cada alvéolo existe um tamanho em que a tensão superficial diminui suficientemente rápido que a pressão começa cair ao invés de continuar a decrescer, e isto faz os alvéolos estabilizarem em cerca de um quarto do seu tamanho máximo. Alvéolos não cobertos com surfactante, tais como aqueles das crianças com RDS, desaba como pequenas bolhas, e uma pressão bem grande é necessária para reabri-los. Uma criança com RDS não deve ter energia para respirar com sua baixa complacência dos pulmões.
As curvas P-V para um pulmão humano extirpado são mostradas na Figura 7.18. Se o pulmão cai completamente em colapso, uma pressão considerável é necessária para começar sua inflação, semelhante ao esforço extra necessário para começar inflar um balão de borracha. Deste ponto o pulmão infla mais facilmente até ele fechar para o seu tamanho máximo. A curva de pressão durante a deflação parece bem diferente. Quando a pressão cair a zero os pulmões ainda retém algum ar. Muito menos pressão é necessária para então tornar a inflar os pulmões, embora a reinflação não segue a curva de deflação. Um processo cíclico em que diferentes curvas são seguidas nas duas metades do ciclo é dito mostrar histerese. A área dentro do laço é proporcional à energia perdida como calor durante o ciclo.

Durante a respiração tida normal o laço de histerese é bem pequeno, como a curva a na curva P-V normal na Fig. 7.19. Se a respiração tida continua sem variar alguns dos alvéolos caem em colapso, e o laço de histerese torna-se ligeiramente maior e desloca para cima para pressões maiores como mostrado pela curva b. Uma respiração profunda reabre o alvéolo, e a curva desloca de volta para a. Tomamos uma tal respiração profunda ocasionalmente sem estar ciente dela( um suspiro). Durante a cirurgia um anestesista ocasionalmente forçará um grande volume de gás para os pulmões do paciente para reabrir os alvéolos que caíram em colapso. Batidas no peito evita um paciente de tomar uma respiração profunda e alguns de seus espaços pulmonares é como se estivesse perdido pelo colapso do alvéolo, ou atelectasia.
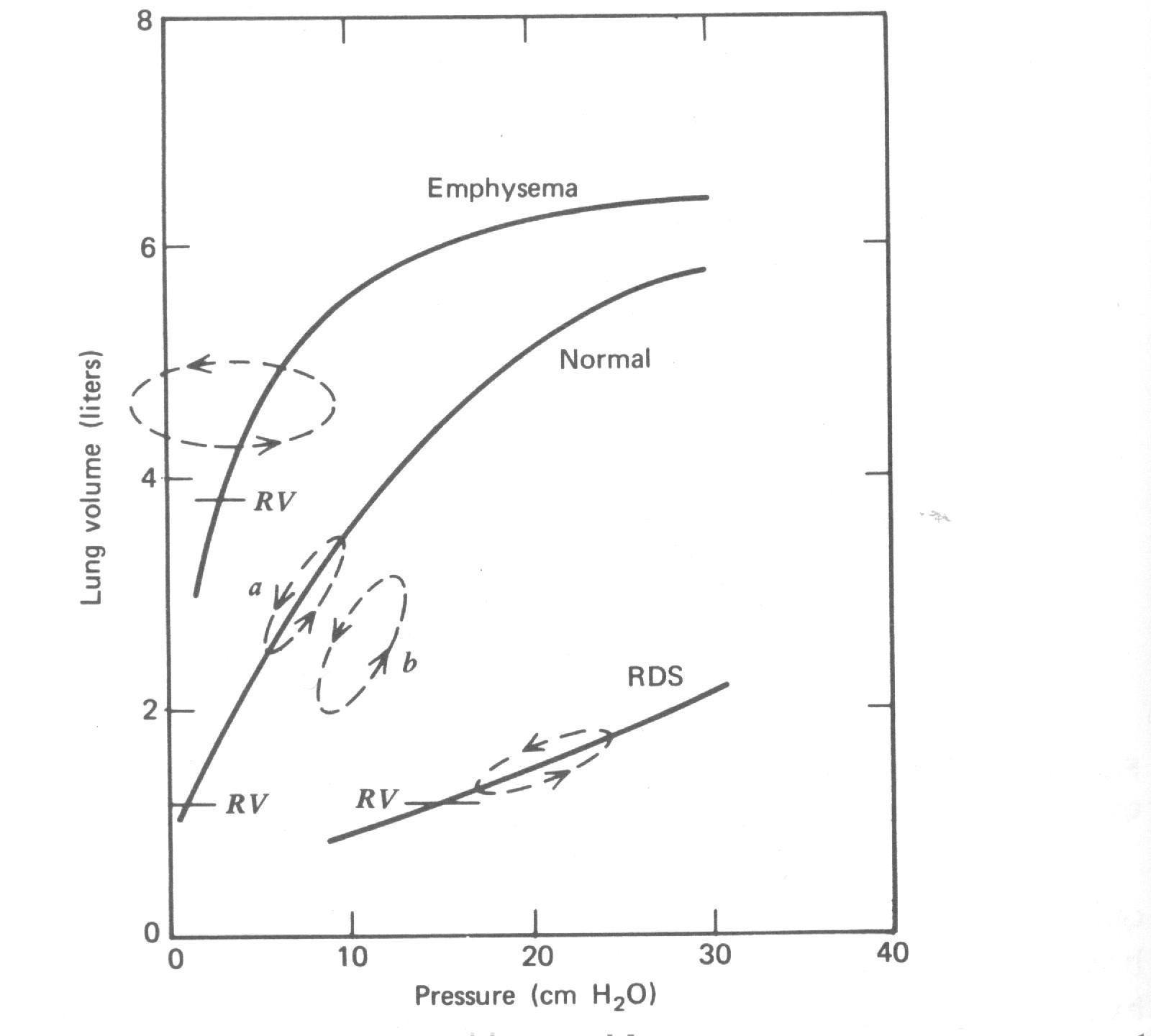
A curva P-V para um pulmão sofrendo de uma falta de surfactante (RDS) é também mostrada na Fig. 7.19; o laço de histerese é deslocado para a direita e uma grande pressão deve ser mantida para segurar os pulmões inflados. Note que a baixa complacência (DV/DP) deste pulmão. A curva P-V de um paciente com enfisema severo é também mostrado na Fig. 7.19; note que o aumento da complacência, o maior volume residual, e a grande área dentro do laço de histerese.
7.6 O MECANISMO DA RESPIRAÇÃO
A respiração está normalmente sob controle inconsciente. Embora a razão de respiração possa ser variada a vontade, uma pessoa é inconsciente de sua respiração a maior parte do tempo a menos que ela esteja sofrendo de asma ou enfisema. O controle fisiológico da respiração depende de muitos fatores, mas o pH no centro respiratório do cérebro exerce o principal controle.
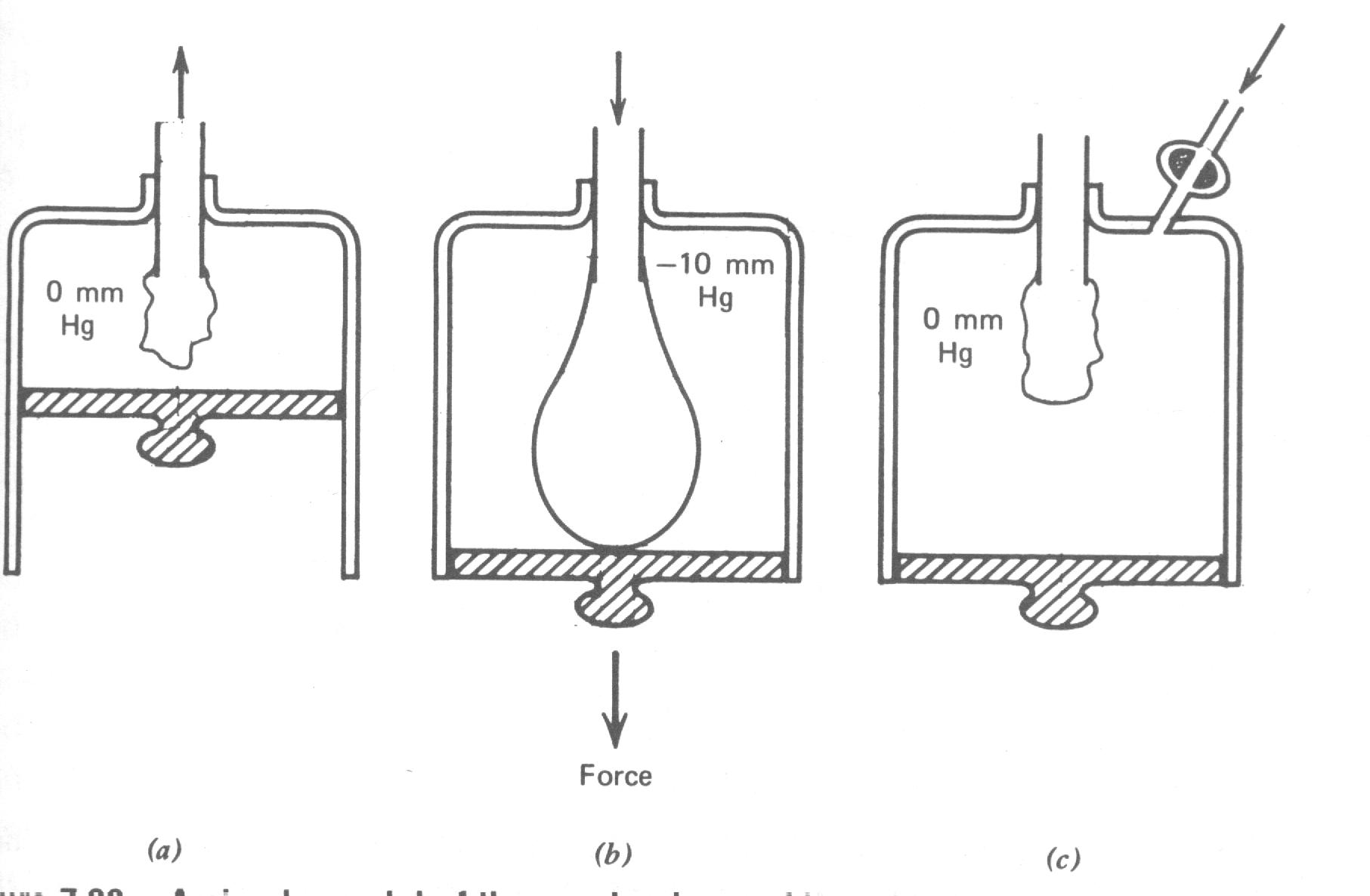
Se um pulmão fosse removido do peito todo o ar seria do peito todo o ar seria escaparia dele e ele murcharia para cerca de um terço do seu tamanho tal qual um balão murcha quando o ar do seu interior vaza. Os pulmões podem ser pensados como milhões de pequenos balões, todos tentando murcharem. Os pulmões normalmente não murcham porque eles estão num recipiente impermeável ao ar – o peito. Quando o diafragma e as costelas movem os pulmões permanecem em contato com eles. Duas forças protegem os pulmões do colapso: (1) a tensão superficial entre os pulmões e a parede do peito e (2) a pressão do ar dentro dos pulmões. A força de tensão superficial é semelhante aquela entre dois pedaços de celofane ou saran wrap stuck together. Se os pulmões dominam esta força e empurram o peito para frente um vácuo poderia ser criado desde que o ar não pode atingir o espaço interpleural. Como o ar dentro dos pulmões está à pressão atmosférica (~ 105 N/m2) ele puxaria os pulmões para trás em contato com as paredes do peito. Existe normalmente uma pressão negativa de 5 a 10 mmHg no espaço interpleural.
Vários músculos estão envolvidos na respiração. Músculos intercostais na parede do peito fazem o peito expandir quando eles contraem. Outros músculos entre o pescoço e tórax podem também contraírem para expandir o peito. Normalmente a maioria da respiração é feita por contração dos músculos do diafragma; estes puxam o diafragma para baixo, expandindo os pulmões. Quando inspiramos, puxamos o diafragma para baixo como mostrado esquematicamente pela flecha na Fig. 7.20b. Isto produz uma pequena pressão negativa nos pulmões e o ar flui para dentro. Quando expiramos , relaxamos os músculos do diafragma, a força elástica nos pulmões fazem o diafragma retornar a sua posição neutra, e o ar flui para fora dos pulmões sem qualquer esforço muscular ativo. Se os músculos do diafragma são paralisados os músculos na parede do peito são usados para respiração.
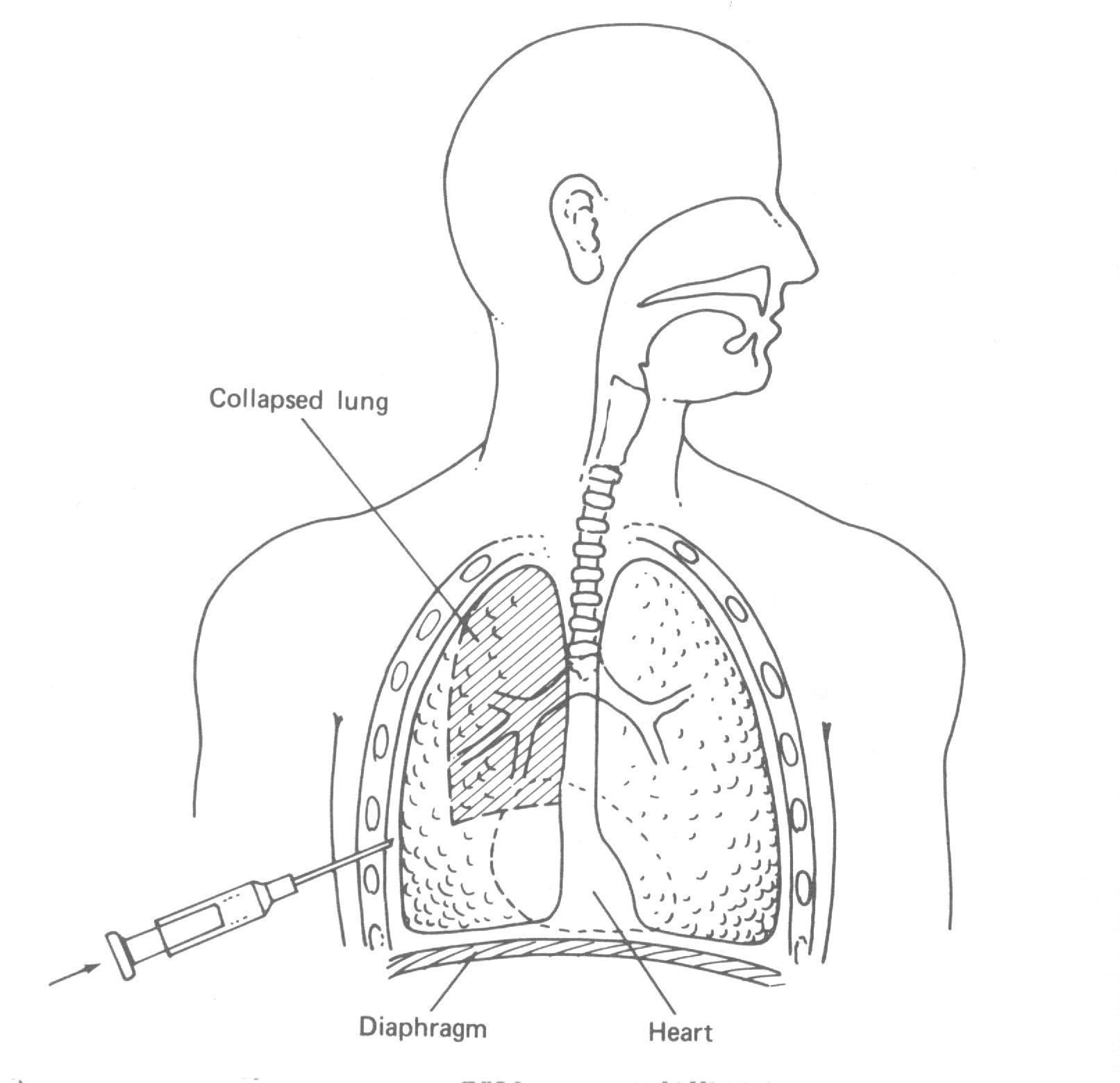
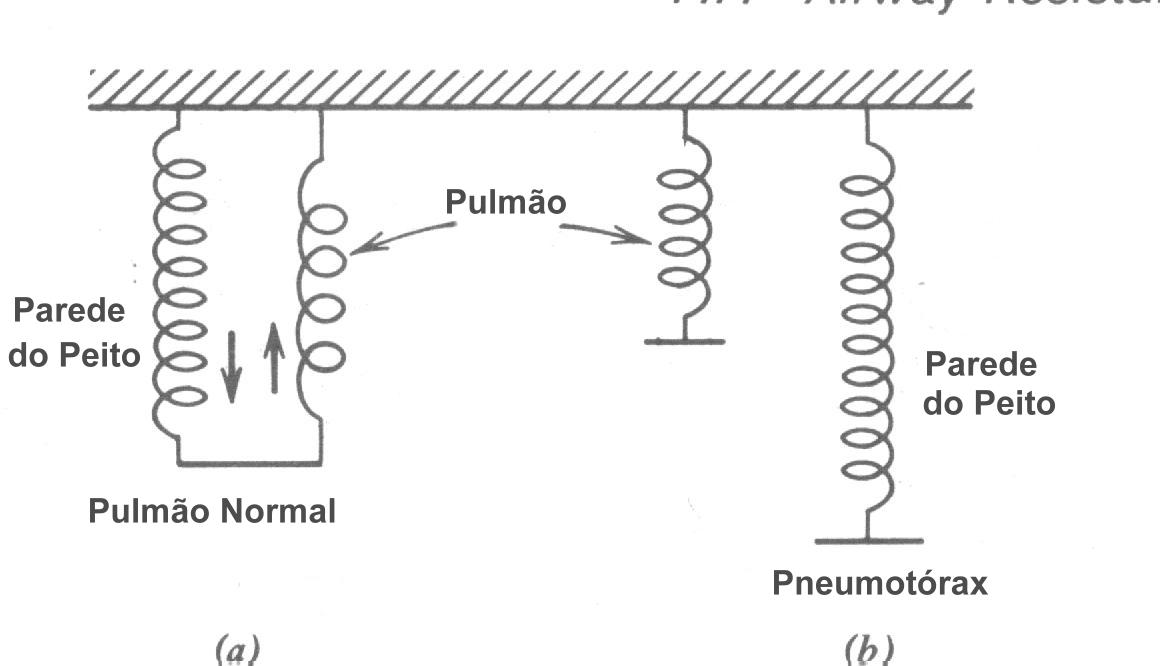
Se a parede do peito é furada como mostra esquematicamente pela válvula abertas na Fig. 7.20c o pulmão colapsa, o diafragma abaixa e a parede do peito expande. Esta condição é conhecida como um pneumotórax (literalmente, ar-peito). Ocasionalmente ela é medicamente desejável colapsar um dos pulmões para permitir ele “descansar”. Como cada pulmão tem seu próprio compartimento lacrado é possível colapsar um pulmão somente como mostrado na Fig. 7.21. Isto é feito mais simplesmente inserindo uma agulha oca entre as costelas (acupuntura intercostal) e permitir o ar fluir para dentro do espaço intratorácico. O ar injetado no espaço é gradualmente absorvido pelos tecidos, e aos pulmões expande ao normal num período de poucas semanas. Algumas vezes um pulmão colapsa espontaneamente devido a nenhuma causa conhecida. Esta condição de pneumotórax espontâneo é moderadamente comum nos estudantes de idade de colegial. Como nos procedimentos médicos, os pulmões retornam ao normal quando o ar é absorvido para os tecidos vizinhos.
Já que os pulmões e o peito são elásticos, podemos representá-los por molas (Fig. 7.22). Sob condições normais eles são acopladas: as molas “pulmonares” são esticadas e as molas “peitorais” são comprimidas (Fig. 7.22a). Durante um pneumotórax, os pulmões e o peito são independentes e as molas representam indo para as suas posições relaxadas como indicada na Fig. 7.22b. Os pulmões colapsam, e as paredes do peito alargam.
O espaço intratorácico nem sempre está numa pressão negativa . Se você fechar seu windpipe e tentar expirar forçadamente, a pressão intratorácica pode tornar-se bem alta. Isto é chamado um manobra de Valsalva. Uma pessoa faz isto quando ela sopra um balão de borracha. Sob mais condições fisiológicas ela faz isto apenas antes de engolir ou espirrar e durante o stress de defecação ou vômito. Aumentando a pressão no peito comprime a principal veia (veia cava) levando sangue de volta ao coração e reduz o volume do sangue bombeado pelo coração. A pressão negativa normal no peito ajuda manter a veia cava aberta. A pressão sangüínea na veia cava é somente 0,5 cm H2O perto do coração.
7.7 - RESISTÊNCIA DAS VIAS AÉREAS
Podemos inspirar mais rapidamente do que podemos expirar. Durante a inspiração a força nas vias aéreas tendem a abri-las mais; durante a expiração as forças tendem a fechar as vias aéreas e restringir assim o fluxo de ar. Para um dado volume pulmonar, a razão do fluxo expiratório atinge um máximo e permanece constante; ela poderia mesmo diminuir um pouco com o aumento da força expiratório.
Pacientes com doenças de obstrução das vias aéreas tais como asma e enfisema acham que um aumento do esforço para expirar diminui a razão de fluxo consideravelmente. Estes pacientes inconscientemente encontram algum alívio mantendo uma grande quantidade de ar nos pulmões, assim mantendo suas vias aéreas tão grandes quanto possível. Eles podem freqüentemente inspirar próximo da razão normal de modo que eles inspiram rapidamente para permitir mais tempo para a expiração. A física pulmonar do enfisema é discutida na Seção 7.9.
O fluxo de ar nos pulmões é análogo ao fluxo de corrente num circuito elétrico. A “lei de Ohm” para o fluxo de ar se parece com a lei de Ohm para circuitos elétricos, com a voltagem trocada por diferença de pressão DP e a corrente trocada pela razão do fluxo de ar DV/Dt ou V. A resistência das vias aéreas Rg é a razão do DP pelo V. A resistência das vias aéreas é dada em unidades de pressão por unidade de razão de fluxo, comumente cm H2O / (litro/s). Nos adultos típicos Rg = 3.3 cm H2O (litros / s). O Rg depende das dimensões do tubo e da viscosidade do gás. A situação é complicada pela complexidade das vias aéreas. A maioria da resistência está na passagem superior das vias aéreas. A área nasal toma cerca da metade do Rg, e outros 20% são devidos às outras vias aéreas superiores. Nos sujeitos normais menos que 10% do Rg está nas vias aéreas terminais. Assim a doença que afetam as vias aéreas terminais (bronquíolos e alvéolos) não afetam apreciavelmente a resistência das vias aéreas até estarem bem avançadas.
A constante de tempo dos pulmões está relacionada à resistência das vias aéreas Rg e a complacência C. Recorde-se que a complacência é DV / DP ( Seção 7.4). O produto RgC é a constante de tempo para os pulmões. Isto é análogo à constante de tempo RC’ para um capacitor C’ descarregar através de uma resistência R num circuito elétrico. A constante de tempo dos pulmões é complicada desde que muitas partes dos pulmões estão interconectadas. Se uma parte do pulmão tem uma constante de tempo maior que as outras partes, ela não obterá sua porção de ar e aquela parte dos pulmões será pobremente ventilada.
7.8 - O TRABALHO DA RESPIRAÇÃO
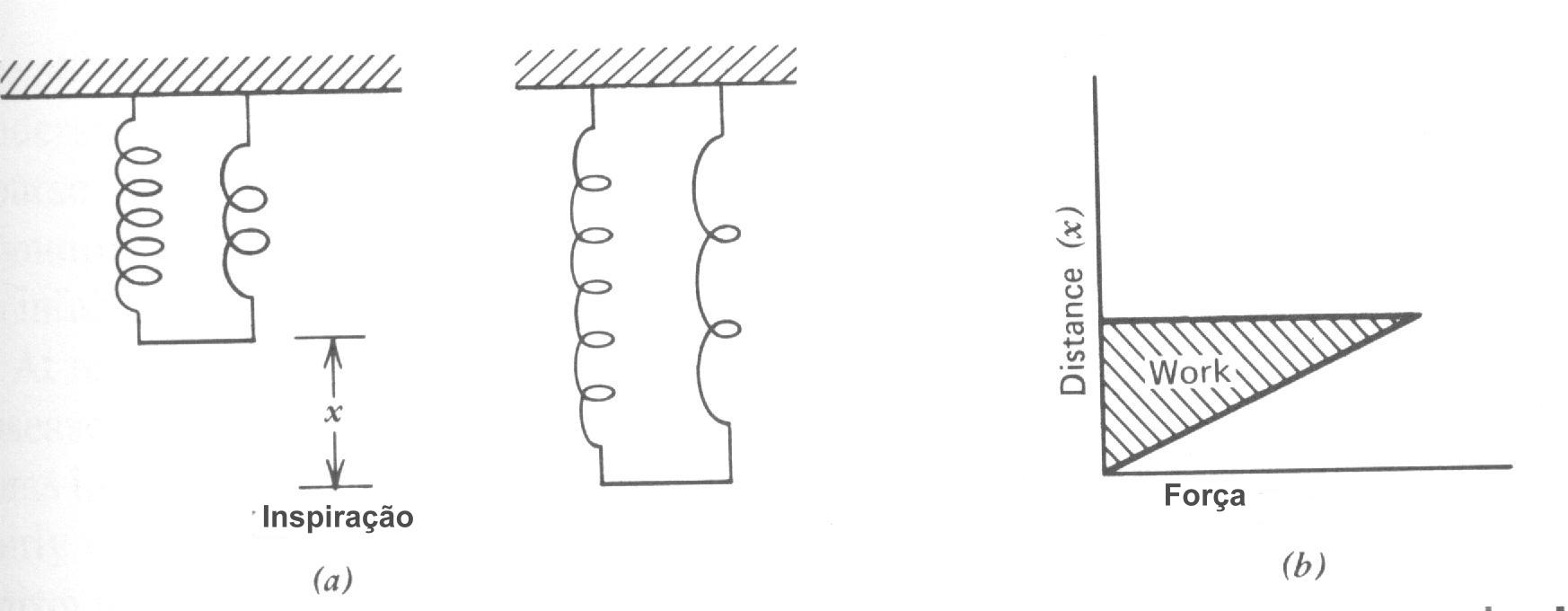
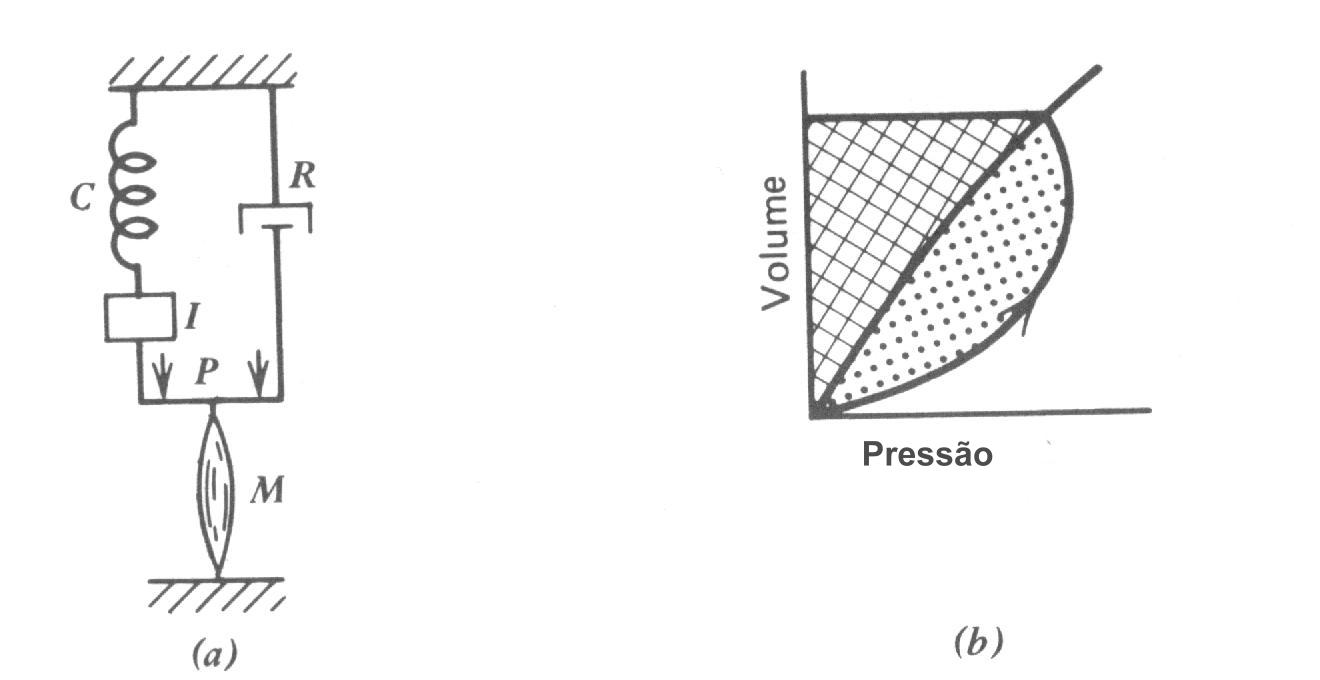
A quantidade de trabalho feito na respiração normal considera para uma pequena fração da energia total consumida pelo corpo (~ 2% no repouso). O principal trabalho da respiração pode ser pensado como o trabalho feito no alongamento da molas representando o sistema pulmão-parede do peito-diafragma (Fig. 7.23a), que é proporcional à área hachuriada na Fig. 7.23b; entretanto, isto é uma super simplificação do trabalho da respiração. Um modelo melhor está mostrado na Fig. 7.24. A resistência dos tecidos e a resistência do fluxo de gás produz calor; este pode ser representado como um freio (R). A elasticidade do sistema peito-pulmão é representada pela mola C. A inércia I da massa dos pulmões e paredes do peito devem também ser superada; na razão de respiração normal, a inércia pode ser desprezada, mas na máxima razão de respiração ( mais de 100 respirações / minutos) ela é um fator significante. O trabalho da respiração está mostrado pela área sombreada total na Fig. 7.24b; A área sombreada mais escura representa o trabalho contra a mola C e a área mais clara representa o trabalho contra a resistência. Durante a respiração normal, nenhum trabalho é feito durante a expiração; os músculos relaxam e as molas “restabelecem de súbito ”para expelir o ar, dissipando a energia no freio R. Durante exercícios vigorosos os músculos são usados para expelir o ar. O trabalho de respiração durante exercícios pesados podem aumentar até 2% o consumo de energia total do corpo.
Respiração rápida e rasa e respiração profunda lenta são ambas menos eficiente que a razão normal. A maioria dos animais ajustam suas razões de respiração no repouso ao uso mínimo de potência. Em baixas razões de respiração a maioria do trabalho é feita contra as forças elásticas do pulmão e peito (área mais escura na Fig. 7.24b); nas razões de respiração rápida o trabalho contra as forças resistivas (área mais clara na Fig. 7.24b) aumenta.
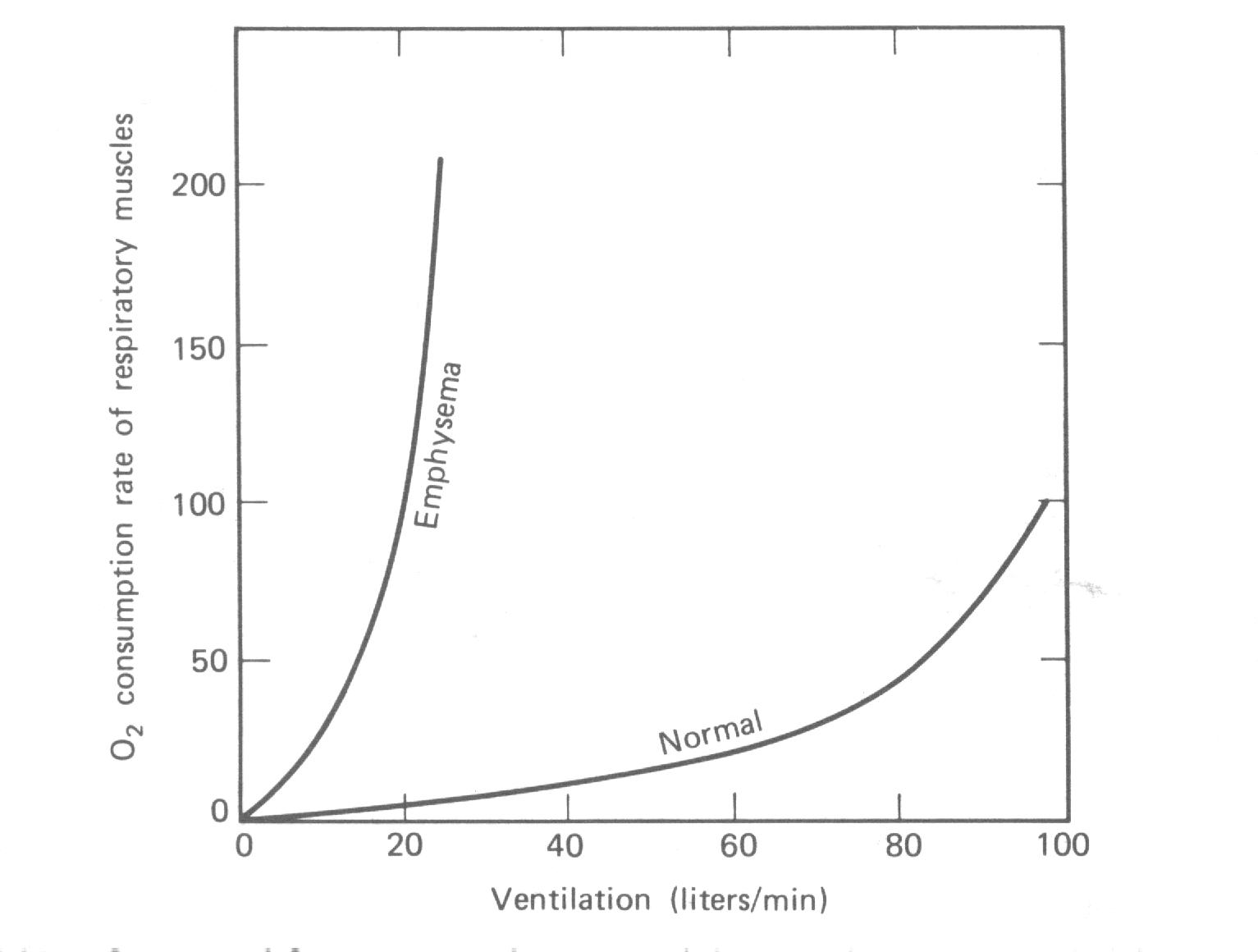
Um outro modo de determinar o trabalho feito na respiração é medir o consumo extra de O2 quando a razão de respiração é aumentada sob condições de repouso. A quantidade de O2 consumido está diretamente relacionada às calorias dos alimentos “queimados”(ver Capítulo 5). Assumimos que o O2 adicional é usado nos músculos respiratórios. A Fig. 7.25 mostra uma curva típica para um sujeito normal e a curva para um paciente com um severo enfisema. O último pode usar mais O2 no trabalho de respiração numa razão mais rápida do que é fornecida pelo seu aumento de ventilação; a quantidade de O2 na sua circulação geral assim cai.
Se compararmos a energia usada na respiração obtida pelo método de consumo de O2 para calcular o trabalho feito usando o modelo mostrado na Fig. 7.24, podemos estimar a eficiência do mecanismo de respiração. Devido as muitas incertezas, a eficiência estima range de 5 a 10%.
A FÍSICA DE ALGUMAS DOENÇAS PULMONARES COMUNS.
As doenças dos pulmões ocorrem numa grande porcentagem de problemas médicos do homem. Estima-se que 15% das pessoas nos U.S.A. na idade de 40 anos tem doença pulmonar detectada. Muitas destas doenças podem ser entendidas em termos de variações físicas nos pulmões. Isto não significa, é claro, que um físico possa curá-las. Os aspectos físicos de algumas doenças comuns dos pulmões são discutidos nesta seção. A física do RDS em crianças está discutida na Seção 7.5.
Em repouso, somente uma pequena fração da capacidade dos pulmões é usada. Assim, uma doença pulmonar que reduz a capacidade freqüentemente não produz um notável sintoma nos seus estágios iniciais. Quando os sintomas são notáveis, a doença está muito bem avançada. Muitos testes das funções pulmonares força o mecanismo da respiração ao seu limite e assim permite detectar variações que não são ordinariamente aparentes. Existem alguns testes pulmonares simples que seriam incluídos em todos os “check up” da saúde.
No enfisema as divisões entre os alvéolos se rompem, produzindo espaços pulmonares maiores. Esta destruição dos tecidos dos pulmões reduz a elasticidade dos pulmões. Os pulmões tornam-se mais complacentes – uma pequena variação na pressão produz uma variação maior do que o normal no volume. Enquanto a primeira vista isto parece tornar mais fácil a respiração, o oposto é verdadeiro. Muito do trabalho da respiração é feito superando a resistência das vias aéreas. No enfisema a resistência das vias aéreas aumenta grandemente.
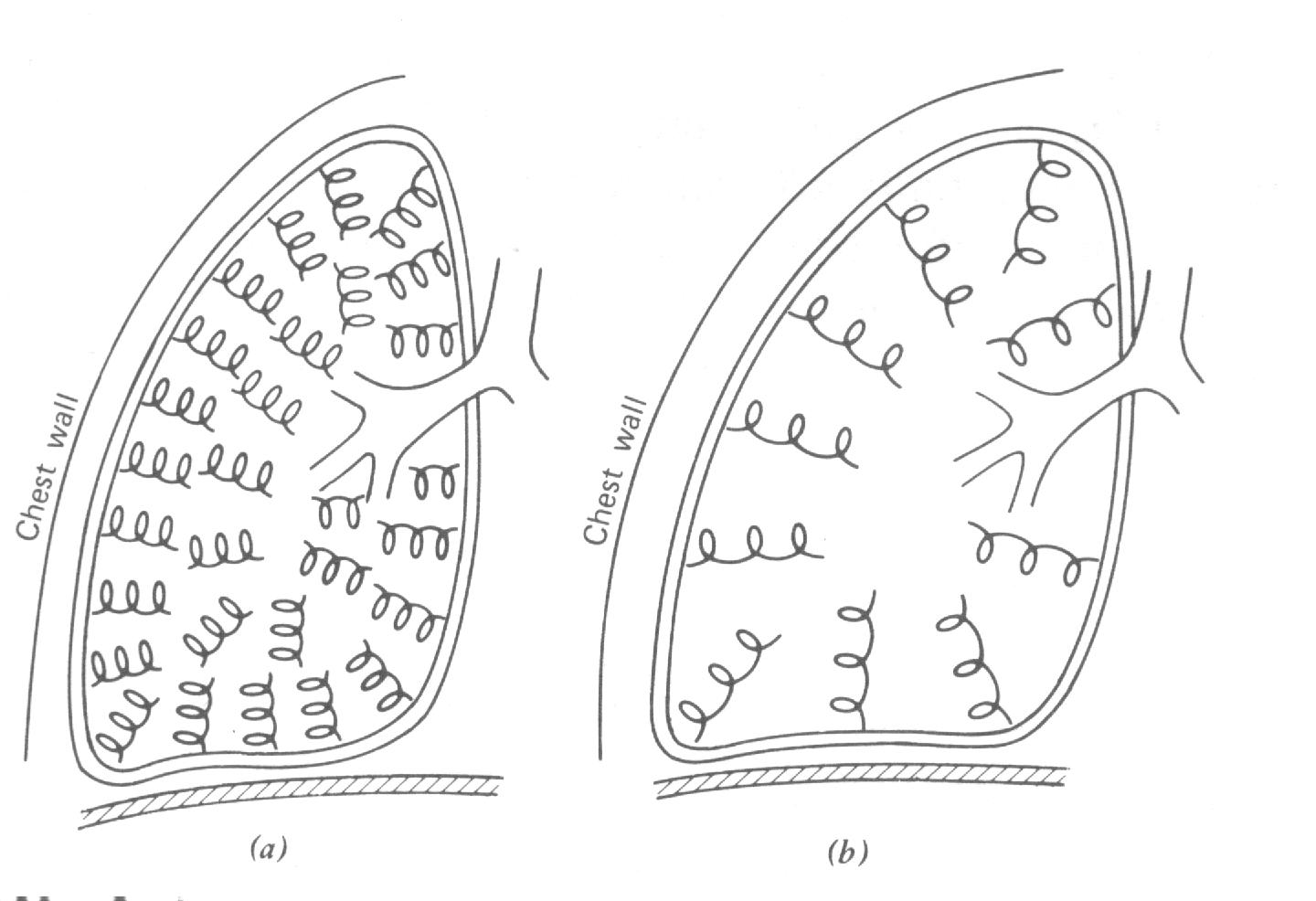
A Fig. 7.26 ajudará você entender a física do enfisema. Você pode pensar na elasticidade dos tecidos no pulmão normal como milhões de pequenas molas interconectadas. Fig. (7.26a). Estas “molas” tendem a murcharem os pulmões e produzirem a força que puxa a parede do peito. Existem também puxamento das paredes das vias aéreas, isto mantém as vias aéreas abertas e ajudam a reduzir a resistência das vias aéreas durante a expiração.
A situação no enfisema severo é mostrada na Fig. 7.26b. O número de “molas” trabalhando foi grandemente reduzido, e aquelas presentes são muito mais fracas que o normal. Isto produz duas importantes variações: (1) O pulmão torna-se frouxo e expande com uma tensão reduzida e permite a parede do peito expandir quase para o volume de repouso da parede do peito sem o pulmão – cerca de 60% da capacidade vital (Fig. 7.12); e (2) os tecidos não puxam muito forte as vias aéreas, permitindo as vias aéreas apertadas murcharem facilmente durante a expiração. Este aumento na resistência das vias aéreas é o principal sintoma de enfisema severo. O aumento do tamanho dos pulmões aumenta a FRC e o volume residual (Fig 7.19). O peito é super inflado e a postura é afetada: alguém com a doença parece peitudo. Como uma pessoa que tem enfisema é incapaz de soprar uma chama, é simples testar esta doença. Enfisema ocorre ocasionalmente nos não fumantes, mas o grande crescimento recente da doença tem sido principalmente entre os fumantes.
Na asma, uma outra doença de obstrução comum, o problema básico é também a dificuldade expiratória devido ao aumento da resistência nas vias aéreas. Algumas destas resistências é aparentemente devido ao inchaço (edema) e muco nas vias aéreas menores, mas muito disto é devido a contração do músculo lisos ao redor das grandes vias aéreas. Complacência pulmonar é essencialmente normal, mas o FRC pode ser maior que o normal desde que o paciente freqüentemente começa inspirar antes de completar a expiração normal.
Na fibrose dos pulmões as membranas entre os alvéolos engrossam. Isto tem dois efeitos marcantes: (1) a complacência dos pulmões decresce e (2) a difusão de O2 nos capilares pulmonares diminui. A resistência expiratória é essencialmente normal. Uma pessoa com a doença terá dores e mesmo dificuldade respiratória (dispnéia) ou deficiência de respiração durante exercícios. Fibrose dos pulmões podem ocorrer se os pulmões tiverem sido irradiados (p.ex, no tratamento do câncer), embora não seja a única causa.
PROBLEMA 7.12 - Num paciente com enfisema severo, qual das seguintes são
BIBLIOGRAFIAJ. M., E. Agostoni, and J. N. Davis, The Respiratory Muscles, Mechanics and Neural Control, 2nd ed., Saunders, Philadelphia, 1970.
Surface Tension en the Lungs, Freeman, San Francisco, 1962.
Physics for Anaesthetics, Thomas, Springfield, Ill., 1976
The Chronic Bronchitis and Emphysema Handbook. John Wiley & Sons, New York, 1990
Human Respiration: A Programmed Course, Freeman, San Francisco, 1968.
Applied respiratory Physiology with Special Reference to Anesthesia, Appleton-Century-Crofts, New York, 1969.
Enjoying Life with Emphysema, Lea and Febiger, Philadelphia, 1984.
Breath Tests in Medicine, Scientific American, 267:74-79 (1992).
Emphysema and Common Sense, Parker Publishing, New York, 1968.
Physiology and Biophysics, 19th ed., Saunders, Philadelphia, 1965, Chapters 39 to 42.
The Clinical Physics and Physiology of Chronic Lung Disease, Inhalation Therapy, and Pulmonary Function Testing, C.C. Thomas, Springfield, IL, 1973
QUESTÕES DE REVISÃO
Esta porcentagem relativamente alta de O2 é a razão a respiração boca a boca é prática e que soprando uma fogueira ajuda a fazer mais brilho (mantê-la acesa).
É assumido que o ar inspirado é seco e o ar expirado é saturado, pH2O = 6,2 kPa.
N.T. – termo adaptado, “que age na superfície”.
Idiopatico é uma palavra útil – ela significa de origem desconhecida.